IMAT Cell Division: Mitosis & Meiosis
🧬 0. Introduction: Genomes, Chromatin, and Chromosomes
The ability of organisms to reproduce their own kind is the one characteristic that best distinguishes living things from nonliving matter. The continuity of life is based entirely on the reproduction of cells, or Cell Division. Before a cell can divide, it must flawlessly organize, duplicate, and segregate its massive library of genetic information.
The Organization of Eukaryotic DNA
- Genome: A cell's total endowment of DNA. In prokaryotes, it is typically a single circular DNA molecule located in the nucleoid region. In eukaryotes, the genome is vastly larger and consists of multiple discrete linear DNA molecules enclosed within a double-membraned nucleus.
- Chromatin: Eukaryotic chromosomes are composed of chromatin, a highly complex, dynamic structural array of DNA intimately associated with specific proteins. During interphase (non-dividing states), chromatin exists as a diffuse, tangled mass.
- Euchromatin: Loosely packed, lightly staining DNA. This region is transcriptionally active, meaning RNA polymerase can access the genes to transcribe mRNA.
- Heterochromatin: Tightly packed, heavily staining, highly condensed DNA. This region is transcriptionally inactive. Centromeres and telomeres are largely composed of constitutive heterochromatin.
- Histones & Nucleosomes: To fit roughly 2 meters of human DNA into a nucleus that is only about $10 \mu m$ in diameter, it must be massively compacted. The DNA double helix wraps exactly twice around a core octamer of 8 basic histone proteins (two each of H2A, H2B, H3, H4) to form a Nucleosome (often described as "beads on a string"). A fifth histone protein (H1) sits outside the core and clamps the DNA to the nucleosome, facilitating higher-order packing into the 30-nm fiber.
- Chromosomes: Strictly prior to cell division, the chromatin supercoils, folds, and condenses massively to form distinct, heavily visible bodies called chromosomes. This extreme physical condensation prevents the delicate, incredibly long DNA strands from tangling, knotting, and breaking during the violent mechanical movements of cellular division.
Somatic cells (all general body cells except the gametes) contain two full sets of chromosomes, one inherited from each parent, making them Diploid (2n). In humans, the diploid number is 2n = 46 (comprising 22 pairs of autosomes and 1 pair of sex chromosomes). Gametes (sperm and eggs) contain only a single, unique set of chromosomes, making them Haploid (n). In humans, n = 23.
Image 1: Hierarchical Organization of Eukaryotic DNA

Visual Analysis: DNA Organization
This visual maps the incredible structural compaction required to fit the genome into the nucleus (LO 11.0).
- Nucleosome Level: Shows DNA wrapping twice around the histone octamer (H2A, H2B, H3, H4) with H1 as the linker.
- Chromatin Fibers: The 30-nm solenoid structure and subsequent looping onto non-histone protein scaffolds.
- Metaphase Condensation: The ultimate supercoiling into the familiar X-shaped chromosome, preventing mechanical damage during division.
Anatomy of a Duplicated Chromosome
Following DNA replication during the S phase, each chromosome consists of two absolutely identical copies. These are formally called Sister Chromatids. They are initially attached firmly together all along their lengths by ring-like protein complexes called Cohesins. They are most tightly and visibly bound at a highly constricted region of DNA called the Centromere. Once the sister chromatids separate during anaphase of mitosis (or anaphase II of meiosis), they are no longer considered chromatids; they are immediately considered full, individual, independent chromosomes.

🔄 1. The Eukaryotic Cell Cycle & Strict Regulation
The cell cycle is the highly ordered, strictly regulated sequence of events from the exact moment a cell is created by the division of a parent cell until its own division into two new cells. It consists of a long, metabolically active preparative growth phase called Interphase and a short, dramatic, physical division phase called the M Phase.
The Cell Cycle Schematic & Checkpoints
Interphase (Accounts for 90% of the Cell Cycle)
- G₁ Phase (First Gap): The cell recovers from previous division, grows physically much larger, copies essential organelles (like mitochondria and ribosomes), and synthesizes molecular building blocks. It carries out its normal physiological metabolic functions.
Crucial Exam Concept: If you observe a normal, functioning cell (like a hepatocyte producing bile or a beta cell secreting insulin), it is currently in the G₁ phase. - S Phase (Synthesis): The cell synthesizes a complete, flawless copy of the nuclear DNA. Chromosomes are duplicated, forming the two sister chromatids. It also duplicates a critical microtubule-organizing structure called the Centrosome (which contains a pair of perpendicular centrioles in animal cells).
- G₂ Phase (Second Gap): The cell grows further, makes proteins and organelles specifically necessary for division (such as massive amounts of tubulin proteins required to build the mitotic spindle apparatus), and begins to reorganize its internal contents in preparation for the onset of mitosis.
The cell cycle is not a passive clock; it is actively driven by a highly complex, exquisitely sensitive chemical control system. The core regulatory molecules are two specific types of proteins: Cyclins and Cyclin-Dependent Kinases (CDKs).
- CDKs are kinase enzymes that are always present in the cell at a relatively constant concentration throughout the entire cycle. However, they are completely inactive on their own.
- Cyclins are regulatory proteins whose intracellular concentration cyclically rises and falls dramatically during specific phases of the cell cycle.
- When a specific Cyclin binds to its corresponding CDK, it creates an active kinase complex. This active complex phosphorylates specific target proteins to physically push the cell forward into the next phase.
Classic Example: MPF (Maturation-Promoting Factor) is the Cyclin B + CDK1 complex that triggers the cell's passage past the G₂ checkpoint directly into M phase by phosphorylating proteins that cause nuclear envelope breakdown and chromosome condensation.
The Three Critical DNA Quality Control Checkpoints:
- 1. The G₁ Checkpoint (The Restriction Point): The most crucial checkpoint in mammalian cells. Checks for adequate cell size, sufficient nutrients, presence of external growth factors, and strictly checks for DNA damage. If a cell receives a "go-ahead" signal here, it is committed to divide and will usually complete the S, G₂, and M phases. If it does not, it exits the cycle entirely, switching into a permanent or semi-permanent nondividing state called the G₀ phase (e.g., mature neurons and cardiac muscle cells). The Rb (Retinoblastoma) protein and p53 act as the master molecular brakes here.
- 2. The G₂ Checkpoint: Checks for any residual DNA damage and rigorously verifies that DNA replication (S phase) is completely finished with high fidelity. Regulated heavily by the accumulation of MPF.
- 3. The M Checkpoint (Spindle Assembly Checkpoint): Occurs midway through mitosis during metaphase. The cell physically pauses division until it can verify that every single chromosome is properly attached to spindle fibers from BOTH opposite poles at their kinetochores. Only when this tension is sensed will the cell activate the Anaphase-Promoting Complex (APC/C) to cleave cohesins.
🧬 2. Mitosis (The M Phase): Asexual Division
Mitosis strictly refers only to the division of the genetic material in the nucleus (Karyokinesis). It is classically broken down into five distinct morphological stages. The ultimate goal is flawless genetic fidelity: producing two genetically identical diploid (2n) daughter cells from a single diploid parent cell.
Image 6: Stages and Mechanics of Mitosis

Visual Analysis: Mitosis Stages
This visual tracks the physical transformation of the cell as it partitions its genome (LO 11.2).
- Morphological Landmarks: Clearly identifies the signature features of Prophase (condensation), Metaphase (alignment), and Anaphase (separation).
- Envelope Dynamics: Shows the fragmentation of the nuclear envelope in prometaphase and its precise reconstruction in telophase.
- Symmetry: Emphasizes the creation of two identical daughter nuclei.
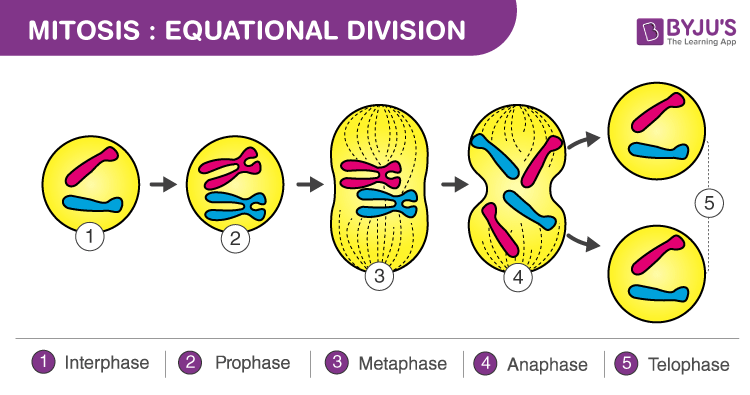
Image 3: Detailed Cellular Mechanics of Mitotic Stages

Visual Analysis: Mitotic Mechanics
A high-resolution look at the molecular engines driving chromosome movement.
- The Spindle Engine: Illustrates the different classes of microtubules (kinetochore, polar, and aster) and their roles in pulling and pushing.
- Kinetochore Attachment: Details the bi-orientation of sister chromatids required for proper segregation.
- Cytokinesis Interface: Shows the coordination between telophase reconstruction and the onset of the actin-myosin contractile ring.
Detailed Cellular Mechanics of Mitotic Stages
The Detailed Sequence of Mitosis
-
Prophase
Condensation & Spindle Assembly: The loosely packed chromatin fibers coil, fold, and condense massively into discrete chromosomes visible under a light microscope. Each duplicated chromosome appears as two identical sister chromatids joined at their centromeres and along their arms by Cohesins. The nucleolus disappears. In the cytoplasm, the mitotic spindle (a massive apparatus composed of microtubules) begins to form between the two separating centrosomes as they migrate to opposite poles of the cell. -
Prometaphase
Nuclear Envelope Breakdown: The nuclear envelope fragments into small vesicles, removing the barrier between the chromosomes and the cytoplasm. The spindle microtubules can now invade the nuclear area. A massive, specialized protein structure called the Kinetochore forms at the centromere of each chromatid. Microtubules attach to these kinetochores (kinetochore microtubules) and begin a violent "tug-of-war", jerking the chromosomes back and forth. -
Metaphase
Alignment and Tension: This is generally the longest stage of mitosis. The centrosomes are now securely anchored at absolute opposite poles of the cell. The chromosomes have been pulled by the opposing forces of the kinetochore microtubules until their centromeres align perfectly on the Metaphase Plate (an imaginary plane equidistant from the two poles).🛑The Spindle Assembly Checkpoint (M-Checkpoint) The cell will physically halt in metaphase until an internal sensor verifies that every single kinetochore is under proper mechanical tension from spindle fibers extending from BOTH opposite poles. If even one chromosome is unattached, the wait signal is maintained, preventing catastrophic aneuploidy. -
Anaphase
Separation & Segregation: The shortest stage of mitosis. It begins suddenly when the APC/C (Anaphase Promoting Complex) unleashes the proteolytic enzyme Separase. Separase violently cleaves the cohesin protein rings holding the sister chromatids together. The two sister chromatids are instantly parted, becoming independent chromosomes.
The liberated chromosomes are rapidly pulled toward opposite poles as their kinetochore microtubules shorten by depolymerizing at their kinetochore ends (a complex "Pac-Man" molecular motor mechanism). Simultaneously, non-kinetochore microtubules extending from opposite poles overlap and push forcefully against each other using motor proteins, elongating the entire cell. -
Telophase
Reconstruction: Two new daughter nuclei form at opposite ends of the cell. Nuclear envelopes are rebuilt from the fragments of the parent cell's original envelope. Nucleoli reappear. The chromosomes become significantly less condensed (uncoiling back into diffuse, functional chromatin). Mitosis—the division of one nucleus into two genetically identical nuclei—is now officially complete.
While mitosis precisely divides the genetic material within the nucleus, cytokinesis physically divides the cell's cytoplasm and organelles. It usually begins concurrently during late anaphase or telophase. Due to structural differences, the mechanism differs completely between animal and plant cells:
Animal Cells (Cleavage)
In animal cells, cytokinesis occurs by a process known as cleavage. A shallow groove called a Cleavage Furrow appears on the cell surface near the old metaphase plate. On the cytoplasmic side of this furrow lies a dynamic contractile ring composed of actin microfilaments associated with myosin motor proteins. The actin and myosin interact, causing the ring to violently contract (analogous to pulling the drawstring of a hoodie) until the parent cell is pinched completely in two, producing two separate daughter cells.
Plant Cells (Cell Plate)
Plant cells possess rigid, tough cellulose cell walls and physically cannot be pinched inward. Instead, during telophase, countless vesicles derived from the Golgi apparatus travel along microtubules to the precise middle of the cell, where they coalesce to form a structure called a Cell Plate. Cell wall materials (pectins and cellulose precursors) carried inside the vesicles collect inside the plate as it grows outward until its surrounding membrane fuses with the plasma membrane, erecting a new cell wall between the daughter cells.
🧬 3. Meiosis: Sexual Reproduction & The Creation of Genetic Variation
Unlike mitosis, which conservatively produces identical diploid clones for growth and tissue repair, Meiosis is designed specifically for sexual reproduction and the propagation of a species. It requires two consecutive cell divisions (Meiosis I and Meiosis II) following only a single initial round of DNA replication. This unique architecture results in four final daughter cells, each containing only half as many chromosomes as the parent cell (a reduction from diploid 2n to haploid n).

Figure 3.1: Reduction and equational divisions of meiosis creating genetic diversity.
Image 3: Phases of Meiosis
Visual Analysis: Meiosis Phases
A comprehensive view of the two-stage division process (LO 11.3).
- Meiosis I (Reductional): Details the pairing and separation of homologous chromosomes, the defining event of sexual reproduction.
- Meiosis II (Equational): Shows the separation of sister chromatids into four genetically unique haploid cells.
- Gametogenesis Link: Visualizes the precursor state for sperm and egg development.
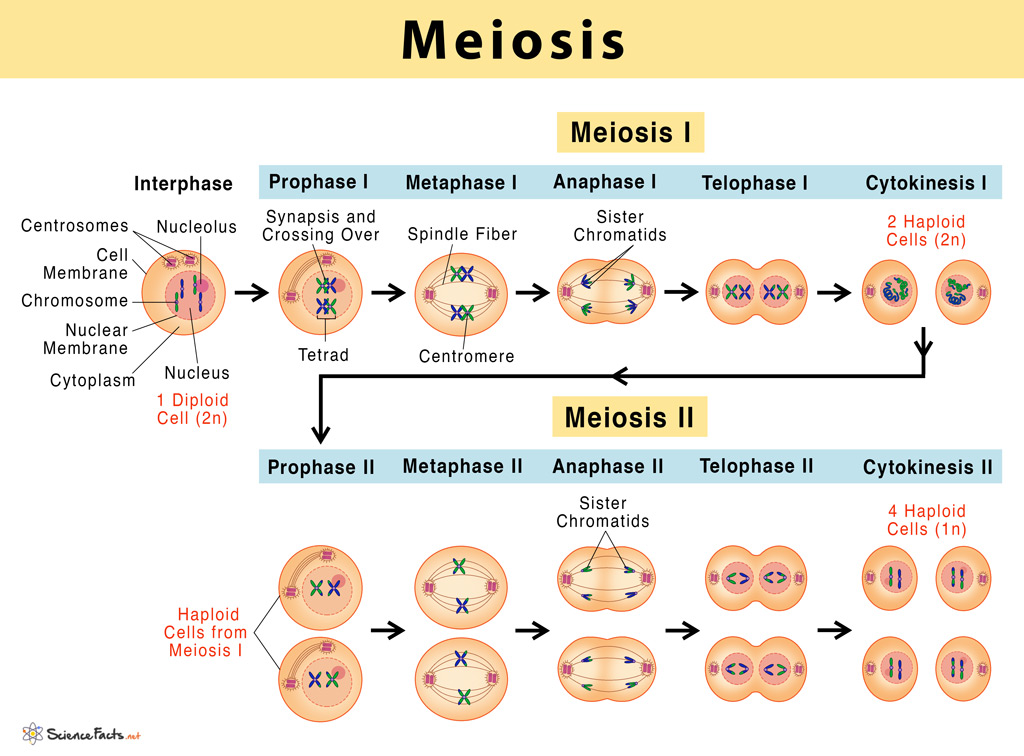
Prophase I: The Engine of Genetic Variation
Prophase I is the longest, most biochemically complex phase of meiosis (in human females, it can pause and last for decades!). It is entirely unique to meiosis and is where the magic of genetic recombination happens. For the IMAT, it is classically divided into 5 distinct morphological sub-stages:
- 1. Leptotene: Chromatin begins to aggressively condense. Chromosomes become visible as thin threads.
- 2. Zygotene: Synapsis begins. Homologous chromosomes (one inherited from the mother, one from the father) find each other and perfectly align gene-by-gene. A zipper-like protein structure called the Synaptonemal Complex physically binds them together tightly along their entire length, forming a Bivalent (or Tetrad).
- 3. Pachytene: Crossing Over occurs. Endonucleases deliberately break the double-stranded DNA of non-sister chromatids. Other enzymes perfectly reattach them to each other, physically exchanging matching segments of DNA. This creates recombinant chromosomes with entirely novel combinations of maternal and paternal alleles. The molecular mechanism involves forming a cross-shaped structure called a Holliday Junction.
- 4. Diplotene: The synaptonemal complex degrades. Homologs attempt to push apart slightly but remain physically tethered together at the exact points where crossing over occurred. These visible X-shaped physical links are called Chiasmata (singular: chiasma).
- 5. Diakinesis: Maximum chromosome condensation is reached. The nuclear envelope breaks down completely, and the spindle apparatus attaches, preparing the tetrads to move to the metaphase plate.
Image 7: Meiosis: Reduction and Variation

Visual Analysis: Reduction & Variation
A high-resolution map of the mechanisms creating genetic diversity.
- Crossing Over: Detail on the formation of chiasmata and the exchange of genetic material between non-sister chromatids.
- Independent Assortment: Shows the random alignment of bivalents at the metaphase I plate, leading to $2^n$ gamete combinations.
- Reductional Logic: Clearly visualizes why and how the chromosome count is halved by the end of Meiosis I.
Meiosis I: Crossing Over & Independent Assortment
Meiosis I: The Reductional Division
Meiosis I is characterized by the separation of homologous chromosomes. This is the definitive stage that halves the chromosome number from diploid (2n) to haploid (n).
- Metaphase I: The homologous pairs (tetrads) align at the metaphase plate. Crucially, their orientation towards the poles is entirely random. This physical phenomenon is the basis for Mendel's Law of Independent Assortment. Because the maternal and paternal homologs can orient facing either pole randomly, there are $2^n$ possible combinations for the resulting gametes. In humans ($n=23$), this alone creates $2^{23}$ (approximately 8.4 million) possible genetic combinations per gamete, even before factoring in the massive variation generated by crossing over!
- Anaphase I: The homologous chromosomes separate and move toward opposite poles. CRITICAL MOLECULAR DISTINCTION: Unlike in mitosis, the cohesin proteins holding the sister chromatids together at the centromere are protected from cleavage by a specialized guardian protein called Shugoshin. Thus, sister chromatids do not separate; they migrate together as a single, combined unit to the same pole. Only the chiasmata are resolved, allowing the homologs to part.
- Telophase I & Cytokinesis: Two haploid cells form. Note that although they are haploid (having only one set of chromosomes), each chromosome still consists of two sister chromatids. There is a brief interphase-like rest period called Interkinesis before Meiosis II begins, but DNA replication DOES NOT occur during this time.
Meiosis II: The Equational Division
Meiosis II is mechanically and morphologically almost identical to Mitosis. The goal is to separate the sister chromatids, which are no longer identical due to the crossing over that occurred in Prophase I.
- Prophase II & Metaphase II: A new spindle apparatus forms, and the chromosomes (still consisting of two chromatids) align individually on the metaphase plate, exactly as they do in mitosis.
- Anaphase II: Shugoshin is degraded, allowing the centromeric cohesins to finally be cleaved by separase. The sister chromatids forcefully separate and are pulled to opposite poles as individual, independent chromosomes.
- Telophase II & Cytokinesis: Nuclei reform around the four sets of chromosomes, which then decondense. The ultimate result is four haploid daughter cells, all genetically distinct from each other and completely distinct from the original parent cell.
Meiosis manifests very differently in the physiology of human males versus females:
Spermatogenesis
Occurs in the seminiferous tubules of the testes continuously from puberty until death. It is a symmetrical, equal division: one primary spermatocyte (2n) yields four functional, equally sized, motile sperm cells (n). The entire process takes roughly 64 days and occurs by the millions every single day.
Oogenesis
Occurs in the ovaries. It is highly asymmetric and heavily interrupted by long pauses.
1. Oogonia enter Meiosis I during fetal development in the womb, but arrest in Prophase I (diplotene stage) and remain frozen until puberty.
2. Following the LH surge at ovulation, Meiosis I completes, but the cytoplasm divides unequally, yielding one large secondary oocyte and one tiny, non-functional Polar Body.
3. The secondary oocyte enters Meiosis II but arrests again in Metaphase II. It only completes Meiosis II if penetration by a sperm physically occurs, yielding a second polar body and one massive, functional ovum.
⚖️ 4. Mitosis vs. Meiosis: The Ultimate IMAT Summary
Image 8: Mitosis vs Meiosis: Summary Table

Visual Analysis: Mitosis vs. Meiosis
A side-by-side comparison of the two fundamental modes of cell division (LO 11.4).
- DNA Replication: Occurs during interphase before both mitosis and meiosis.
- Division Count: Mitosis involves one division, while meiosis requires two successive divisions.
- Synapsis: Occurs only in meiosis, where homologous chromosomes pair and cross over.
- Genetic Outcome: Mitosis preserves diploidy and identity; meiosis creates haploid diversity.
| Property / Feature | Mitosis | Meiosis |
|---|---|---|
| Biological Purpose | Growth, tissue repair, and asexual reproduction. | Production of gametes exclusively for sexual reproduction. |
| Number of Divisions | One | Two |
| Synapsis & Crossing Over? | No. Homologs completely ignore each other. | Yes. Occurs exclusively in Prophase I. |
| Metaphase Alignment | Individual chromosomes align in a single file line. | Meta I: Homologous pairs (tetrads) align. Meta II: Individual chromosomes align. |
| Anaphase Separation | Sister chromatids separate. | Ana I: Homologous chromosomes separate. Ana II: Sister chromatids separate. |
| Final Outcome | 2 identical Diploid (2n) cells | 4 distinct Haploid (n) cells |
🚨 5. Clinical Pathology: Chromosomal Abnormalities & Cancer
The intricate machinery of cell division is generally accurate, but catastrophic errors can and do occur. When these errors happen in meiosis, they are passed to the offspring, leading to severe congenital genetic disorders. When cell cycle regulation fails in somatic cells, it leads to cancer. The IMAT tests heavily on your ability to recognize the mechanisms and clinical presentations of these anomalies.
The Mechanics of Nondisjunction
Nondisjunction is the failure of homologous chromosomes or sister chromatids to separate properly during cell division. The timing of the error dictates the outcome:
Image 5: Mechanics of Nondisjunction

Visual Analysis: Nondisjunction Mechanics
A step-by-step look at the errors that lead to aneuploidy (LO 11.5).
- Meiosis I Error: Homologs fail to separate, resulting in 100% abnormal gametes (n+1 and n-1).
- Meiosis II Error: Sister chromatids fail to separate, resulting in 50% abnormal and 50% normal gametes.
- Clinical consequence: Visualizes the source of trisomies (like Down Syndrome) and monosomies.
Nondisjunction in Meiosis I vs Meiosis II
- Nondisjunction in Meiosis I: Homologous chromosomes fail to separate during Anaphase I. Result: 100% of gametes are abnormal (two are n+1, two are n-1). This is the most common cause of Down Syndrome (associated with maternal age as primary oocytes are paused in Prophase I).
- Nondisjunction in Meiosis II: Sister chromatids fail to separate during Anaphase II in one of the cells. Result: 50% normal gametes, 50% abnormal (one n+1, one n-1).
High-Yield Aneuploidies
- Trisomy 21 (Down Syndrome): Karyotype: 47, XX,+21 or 47, XY,+21. The most common viable trisomy in humans. Clinical features: intellectual disability, epicanthic folds, single palmar crease, congenital heart defects.
- Trisomy 18 (Edwards Syndrome): Severe developmental issues, micrognathia, prominent occiput, rocker-bottom feet, clenched fists with overlapping fingers. Usually fatal within 1 year.
- Trisomy 13 (Patau Syndrome): Severe intellectual disability, microphthalmia (small eyes), cleft lip/palate, polydactyly. Usually fatal within months.
Better tolerated than autosomal aneuploidies due to X-inactivation (formation of Barr bodies) and the small number of genes on the Y chromosome.
- Klinefelter Syndrome (47, XXY): Male phenotype. Hypogonadism, sterile. Gynecomastia, tall stature. One Barr body present.
- Turner Syndrome (45, X0): Female phenotype. The ONLY viable human monosomy. Short stature, webbed neck (cystic hygroma), sterile (streak ovaries). Zero Barr bodies.
Structural Chromosomal Abnormalities
Physical breakage of a chromosome can lead to four types of changes in chromosome structure. These often occur due to errors during crossing over in Prophase I or exposure to severe radiation:

- Deletion: Loss of a chromosomal segment.
IMAT Example: Cri-du-chat syndrome (deletion on the short arm of chromosome 5p). - Duplication: Repetition of a segment.
- Inversion: A segment reverses its orientation completely within the chromosome.
- Translocation: Movement of a segment to a non-homologous chromosome.
IMAT Example: The Philadelphia Chromosome. A reciprocal translocation between chromosomes 9 and 22 ($t(9;22)$). This fuses the BCR gene with the ABL kinase gene, creating a hyperactive mutant kinase that drives unchecked white blood cell division, resulting in Chronic Myelogenous Leukemia (CML).
When the Cell Cycle Fails: Cancer
Cancer is fundamentally a disease of the cell cycle. It occurs when a cell accumulates mutations that allow it to bypass normal regulatory checkpoints (G1, G2, M), ignore density-dependent inhibition, and divide uncontrollably.
Proto-oncogenes are normal cellular genes that code for proteins that stimulate normal cell growth and division. When mutated or overexpressed, they become cancer-causing Oncogenes. A mutation in just one allele is sufficient (dominant effect).
- Ras gene: Codes for a G-protein that relays signals from a growth factor receptor on the plasma membrane to a cascade of protein kinases. A mutated Ras protein is hyperactive and issues signals on its own, leading to excessive division. Found in ~30% of human cancers.
Genes whose normal products inhibit cell division. They repair damaged DNA, control cellular adhesion, and initiate apoptosis if damage is irreparable. Both alleles must be mutated to lose function (recessive effect, "two-hit hypothesis").
- p53 gene: The "Guardian of the Genome". It halts the cell cycle at G1 if DNA is damaged by UV light or toxins. If repair fails, p53 triggers apoptosis. Mutated in >50% of all human cancers.
- Rb gene: The classic gatekeeper of the G1 restriction point. Binds and inhibits E2F transcription factors.
📝 Mastery Practice: IMAT Cell Division Quiz
Test your comprehensive understanding of the advanced cell cycle, mitosis, meiosis, and clinical pathology covered in this module. These questions are meticulously styled after the rigorous, integrated, multi-step logic required for the IMAT examination.