LESSON 23: CELLULAR ARCHITECTURE
& MOLECULAR PHYSIOLOGY
An exhaustive, integrated treatise encompassing Evolutionary Biology, Membrane Thermodynamics, Intracellular Trafficking, and Biological Stasis.
Course Architecture
Module 1: Foundations
Module 2: Membranes
- 2.1 The Container of Life & Boundary Thermodynamics
- 2.2 Amphipathic Nature & The Hydrophobic Effect
- 2.3 Membrane Fluidity, MODs & Lipid Rafts
- 2.4 Biogenesis: From ER Synthesis to Golgi Refinement
- 2.5 Transverse Asymmetry & Apoptotic PS Exposure
- 2.6 Membrane Proteins: Integral, Peripheral & Hydropathy
- 2.7 The Glycocalyx: Recognition, Protection & Lubrication
Module 3: Transport
- 3.1 Fick's Law & The Permeability Hierarchy
- 3.2 Transporters vs Channels: Saturable vs Linear Kinetics
- 3.3 Primary & Secondary Active Transport Mechanisms
- 3.4 Ion Channels: Selectivity Filter & Atomic Mimicry
- 3.5 Membrane Potential Physics & Nernst Equation
- 3.6 Action Potentials & SNARE-mediated Synaptic Transmission
Module 4: Trafficking
- 4.1 Compartmentalization: Microenvironments & Sequestration
- 4.2 Protein Sorting: The SRP Cycle & Co-translational Import
- 4.3 Vesicle Biogenesis: COPI, COPII, & Clathrin Dynamics
- 4.4 Targeting & Fusion: Rab GTPases & The SNARE Winch
- 4.5 Endocytosis: Phago/Pino/Receptor-Mediated & LDL Pathology
- 4.6 Energy Organelles: Mitochondrial Dynamics & Peroxisomes
Module 5: The Nucleus
Module 6: Cytoskeleton
Module 7: Cell Cycle
Module 8: Reproduction
Module 9: Preservation
IV. Master Final Examination
Module 1: Foundations of Cytology
1.1 Introduction to Biology & Thermodynamics
Defining Life through Physics.
The term Biology (derived from the Greek words bios, meaning life, and logos, meaning study or discourse) represents the scientific categorization of entities exhibiting a specific set of complex, emergent properties. However, "life cannot be defined by the presence of a single magic molecule. As physicist Erwin Schrödinger postulated in his seminal 1944 work What is Life?, living organisms are fundamentally Dissipative Structures. They exist far from thermodynamic equilibrium. While the universe universally trends toward maximum entropy (disorder) according to the Second Law of Thermodynamics, biological entities actively consume energy (eating "negative entropy") from their environment to maintain highly ordered internal states.
Gibbs Free Energy Equation
$$\Delta G = \Delta H - T\Delta S$$Biological systems couple exergonic reactions ($\Delta G < 0$, ATP hydrolysis) to endergonic ones ($\Delta G > 0$, building complex structures) to overcome local entropy decreases.
The Six Pillars of Vitality (Autopoiesis)
For an entity to be scientifically classified as a living organism, it must autonomously perform the following six functions, collectively defining the concept of Autopoiesis (self-creation and self-maintenance):
The ability to generate functional offspring and transmit a stable genetic blueprint (DNA/RNA). This blueprint must be subject to random mutation, generating phenotypic variation for natural selection.
The capacity to actively extract free energy from the environment (autotrophy or heterotrophy) to synthesize ATP, driving entropically unfavorable anabolic reactions.
A directed, highly coordinated increase in biological mass and spatial differentiation, strictly orchestrated by the internal genomic program rather than passive accretion.
The ability to detect physical or chemical perturbations (photon gradients, toxins) and execute a rapid, coordinated cellular response to restore homeostasis.
Self-directed mechanical displacement, spanning from the microscopic streaming of organelles along cytoskeletal tracks to whole-organism locomotion.
The systematic isolation and active physical extrusion of toxic metabolic byproducts (e.g., nitrogenous wastes, ROS) to prevent internal cellular necrotic death.
1.2 Evolution & History of Cell Theory
The Genesis of the Cellular Paradigm.
Life on Earth originated approximately 4 billion years ago in the primordial oceans. Every extant organism on the planet is the result of an unbroken, continuous chain of cellular divisions tracing back to the LUCA (Last Universal Common Ancestor). The profound conceptual realization that all biological complexity is fundamentally cellular did not occur overnight; it required centuries of iterative optical engineering and massive theoretical paradigm shifts, culminating in the formalization of Cell Theory.
The Chronology of Cytological Discovery
Using a primitive compound microscope, Hooke observed razor-thin slices of cork (oak bark). He noted distinct, repeating honeycomb-like compartments and coined the term "cellulae" (Latin for small rooms, likening them to monks' quarters). Crucially, he was observing dehydrated, dead cellulose cell walls entirely devoid of living protoplasm.
A Dutch draper and master lens crafter. Through obsessive glass polishing, he created single-lens microscopes of unprecedented magnification (up to 275x). He became the first human to observe living, motile single-celled organisms in pond water and human dental plaque, which he affectionately termed "animalcules".
Scottish botanist who, while studying orchid epidermal cells, noticed an opaque spot in every cell. He recognized it as an essential, ubiquitous cellular component and named it the Nucleus.
Schleiden (studying plant tissue) and Schwann (studying animal cartilage) synthesized years of disparate observations to formally propose the first two monumental tenets of Cell Theory: 1) All living organisms are composed of one or more cells. 2) The cell is the basic structural and functional unit of life.
Virchow published the definitive maxim "Omnis cellula e cellula" (Every cell stems from another pre-existing cell), establishing the third crucial tenet of Cell Theory. Shortly after, Pasteur's elegant swan-neck flask experiments definitively disproved the ancient, prevailing theory of Spontaneous Generation.
Formalized the Endosymbiotic Theory, proposing that eukaryotic organelles (specifically mitochondria and chloroplasts) originated from free-living prokaryotes that were engulfed by a primordial eukaryotic ancestor. This beautifully explained why these organelles possess their own circular DNA and 70S ribosomes, fundamentally altering our understanding of eukaryotic evolution.
1.3 The Scale of Life & Microscopy
Optical physics and resolution limits.
Understanding cytology requires the mind to operate fluently across logarithmic scales, specifically in nanometers ($10^{-9}$ m) and micrometers ($10^{-6}$ m). Visualizing these structures is fundamentally bounded by the physical properties of the electromagnetic spectrum used to observe them.
The Physics of Resolution (Abbe's Limit)
The resolution ($d$, the minimum distance at which two distinct points can be distinguished as separate entities) of an optical microscope is strictly limited by the diffraction of light, described by Ernst Abbe's 1873 equation:
- $\lambda$: Wavelength of illumination. Visible light is roughly 400–700 nm.
- $n \sin \theta$: Numerical Aperture (NA) of the objective lens. Maximum practical NA in oil immersion is ~1.4.
- Conclusion: The absolute theoretical limit for a light microscope is $d \approx 200 \text{ nm}$. Therefore, to observe viruses, ribosomes, or individual lipid bilayers, biologists must employ an Electron Microscope (EM). By using an electron beam with an exceptionally short wavelength (often $< 0.01 \text{ nm}$), EMs can achieve resolutions down to $\sim 0.1 \text{ nm}$ (atomic scale).
The Logarithmic Scale of Biology
| Biological Object | Approximate Size | Scientific Notation | Required Microscopy |
|---|---|---|---|
| Atom (Hydrogen) | 0.1 nm (1 Å) | $$10^{-10} m$$ | Electron Microscope (TEM) |
| DNA Double Helix (Width) | 2 nm | $$2 \times 10^{-9} m$$ | Electron Microscope (TEM) |
| Plasma Membrane Thickness | 5 nm | $$5 \times 10^{-9} m$$ | Electron Microscope (TEM) |
| Ribosome | 25 - 30 nm | $$3 \times 10^{-8} m$$ | Electron Microscope (TEM) |
| Viruses (e.g., HIV, SARS-CoV-2) | 50 - 150 nm | $$10^{-7} m$$ | Electron Microscope (TEM/SEM) |
| Bacteria / Mitochondria | 1 - 5 µm | $$10^{-6} m$$ | Optical (Light) Microscope |
| Eukaryotic Cells (Animal/Plant) | 10 - 100 µm | $$10^{-5} m$$ | Optical (Light) Microscope |
| Frog Egg / Squid Giant Axon | 1 mm | $$10^{-3} m$$ | Naked Eye |
1.4 Taxonomy, Domains & Viral Classification
Organizing the diversity of life.
The Three Domains of Life (Carl Woese, 1990)
Historically, macroscopic morphology dominated taxonomy (creating the classical 5 Kingdom system). In 1990, Carl Woese radically altered biology by introducing molecular phylogeny. By sequencing and comparing the highly conserved 16S (prokaryotes) and 18S (eukaryotes) ribosomal RNA genes, he proved that life is fundamentally tripartite, revealing that Archaea are genetically and evolutionarily distinct from true Bacteria, despite their similar visual appearance under a microscope.
The "true bacteria (Eubacteria). Ubiquitous in all environments on Earth. Characterized by cell walls strictly composed of cross-linked peptidoglycan. This domain encompasses all known bacterial human pathogens (e.g., Staphylococcus, E. coli) as well as the vast majority of the symbiotic human microbiome.
Prokaryotic in cellular appearance, but their translation and transcription machinery closely mirrors that of eukaryotes. Often extremophiles (halophiles, methanogens, hyperthermophiles). Cell walls utilize pseudomurein instead of peptidoglycan. Their plasma membranes feature unique ether-linked branched isoprenoid lipids to withstand extreme thermal or acidic stress.
Cells possessing a true, double-membrane-bound nucleus and highly compartmentalized internal organelles (Mitochondria, ER, Golgi). Encompasses the classical macroscopic kingdoms: Protista, Fungi, Plantae, and Animalia. Capable of extreme multicellular specialization.
The Viral Exception: Acellular Parasites
Viruses defy the cellular definition of life. They are obligate intracellular parasites. Devoid of ribosomes, mitochondria, and independent metabolic pathways, they exist as inert, crystalline macromolecular complexes (virions) outside a host cell, but transform into highly efficient, replicative machines immediately upon intracellular entry.
Enveloped Virus Model
- The Genome: A virus carries either DNA or RNA, but never both simultaneously. The genome can be single-stranded (ss), double-stranded (ds), linear, or circular.
- The Capsid: A highly ordered geometric protein shell (often mathematically perfect icosahedral or helical shapes) composed of repeating identical subunits (capsomeres) that encases and physically protects the fragile nucleic acid payload.
- The Envelope (Optional): Many significant human pathogens (e.g., HIV, Influenza, SARS-CoV-2) possess an outer lipid bilayer. Because viruses lack lipid synthesis machinery, this envelope is physically stolen from the host cell's plasma membrane or ER during the viral budding process.
- Surface Spikes (Glycoproteins): Protruding from the envelope or capsid, these highly specific proteins dictate viral tropism—binding exclusively to specific host cellular receptors (e.g., HIV gp120 binding to CD4 on helper T-cells).
Viruses are classified into 7 distinct groups based entirely on their genome type (dsDNA, ssDNA, dsRNA, +ssRNA, -ssRNA) and their mandatory pathway to synthesize functional mRNA. For example, Class VI (Retroviruses like HIV) contain +ssRNA but utilize a unique, virally encoded enzyme, Reverse Transcriptase, to convert RNA back into DNA, forcefully violating the central dogma of molecular biology before integrating into the host genome.
1.5 Prokaryotic vs Eukaryotic Architecture
The fundamental structural divide of all life.
The presence or absence of a nuclear envelope is the most profound architectural divide in cellular biology. This structural distinction dictates the mechanisms of gene expression, the logistics of intracellular transport, the magnitude of the cell, and the methodology of cell division. Eukaryotic cells represent a massive leap in complexity, enabling the evolution of multicellular organisms.
Architectural Comparison
| Feature | Prokaryotic (Bacteria/Archaea) | Eukaryotic (Animals/Plants/Fungi) |
|---|---|---|
| Nucleus | Absent. DNA sits naked in an undefined, irregular Nucleoid region in the cytoplasm. | Present. DNA is safely enclosed by a complex, double-membrane nuclear envelope equipped with selective pores. |
| Organelles | None (No membrane-bound compartments). All metabolic reactions (including respiration and photosynthesis) occur freely in the cytoplasm or are attached directly to the plasma membrane. | Extensively compartmentalized. Possesses specialized membrane-bound organelles (Mitochondria, Golgi Apparatus, Endoplasmic Reticulum, Lysosomes, Peroxisomes). |
| DNA Structure | A single, continuous circular chromosome. Haploid (one copy). Naked DNA (not wrapped around histones in bacteria). Extrachromosomal Plasmids are very common and easily exchanged. | Multiple, discrete linear chromosomes. Diploid (homologous pairs in somatic cells). DNA is tightly complexed with basic Histone proteins to form dense, regulated Chromatin. |
| Ribosomes | 70S (composed of a 30S small subunit and a 50S large subunit). Free floating. | 80S (composed of a 40S small subunit and a 60S large subunit). Located free in the cytosol or bound to the membrane of the Rough ER. |
| Cell Wall | Almost universally present. Chemically complex. Made of cross-linked Peptidoglycan in Bacteria and Pseudomurein in Archaea. | Present only in Plants (Cellulose) and Fungi (Chitin). Completely absent in Animal cells, which instead rely entirely on an internal cytoskeleton and extracellular matrix. |
| Cell Division | Binary Fission. Direct replication of the circular chromosome followed by physical splitting. Extremely rapid (e.g., E. coli can divide every 20 minutes under optimal conditions). | A highly regulated, complex process of Mitosis (somatic cells) and Meiosis (gametes) involving a massive microtubule spindle apparatus and strict biochemical checkpoints. |
Clinical Pharmacology: The Principle of Selective Toxicity
The holy grail of antimicrobial chemotherapy is to eradicate the invading bacterial pathogen without causing collateral damage to the human host. This is achieved by rationally designing drugs that exploit the fundamental structural differences between prokaryotic and eukaryotic cells outlined above.
- Inhibitors of Cell Wall Synthesis (e.g., Penicillins, Cephalosporins): Beta-lactam antibiotics irreversibly bind to and inhibit Penicillin-Binding Proteins (PBPs), halting the transpeptidation (cross-linking) of the peptidoglycan wall. Because human cells completely lack a cell wall, they are fundamentally immune to this mechanism. The bacteria, unable to maintain osmotic pressure, lyse and die.
- Inhibitors of Protein Synthesis (e.g., Tetracyclines, Macrolides, Aminoglycosides): These drugs are mathematically designed to bind specifically to the distinct structural clefts of the bacterial 70S ribosome (either the 30S or 50S subunit), halting translation. The human cytosolic 80S ribosome has a different structural conformation and remains completely unaffected. (Clinical Note: Because human mitochondria evolved from bacteria and still possess 70S ribosomes, very high doses of these antibiotics can cause mild mitochondrial toxicity).
1.6 Cell Morphology & Tissue Culture
In Vitro Phenotypes and Immortalization.
When mammalian cells are extracted from the complex 3D architecture of a living tissue and placed into an artificial, 2D plastic petri dish containing nutrient media (In Vitro Cell Culture), they undergo significant morphological adaptation. Their resultant shape is primarily dictated by their embryonic origin and the internal tension generated by their actin cytoskeleton. They are broadly categorized into four phenotypic classifications:
Fibroblast-like
Anchorage-DependentDerived from mesenchymal connective tissue. They exhibit an elongated, bipolar, or multipolar spindle shape. They form strong focal adhesions with the substrate and are highly motile, crawling across the dish using lamellipodia.
Epithelial-like
Anchorage-DependentFlattened, polygonal geometry. They do not crawl independently but grow outwards in discrete patches. Through tight cell-cell junctions (E-cadherins), they form a continuous, impermeable 2D monolayer, reminiscent of pavement stones.
Lymphoblast-like
Suspension CultureDerived from blood or immune tissues (e.g., Leukemias). They utterly lack the integrin machinery to attach to plastic substrates. They grow freely suspended in the liquid media, maintaining a perfectly spherical, uniform morphology.
Neuronal-like
Anchorage-DependentExhibit highly irregular, complex cell body shapes with extremely long, fine branching processes (representing developing axons and dendrites) that attempt to form interconnected synaptic networks across the culture dish.
Experimental Methodology: Primary Culture vs. Immortalized Cell Lines
Primary Cells isolated directly from a donor tissue retain their original, unadulterated physiological functions and genetic stability. However, they are subject to the Hayflick Limit—they will undergo replicative senescence and die after approximately 50 divisions due to the progressive shortening of their telomeres.
Continuous (Immortalized) Cell Lines (such as the famous HeLa cells derived from Henrietta Lacks' cervical carcinoma in 1951) contain severe genetic mutations. Typically, these mutations involve the permanent upregulation of the enzyme Telomerase (which infinitely rebuilds telomeres) and the disabling of critical tumor suppressor genes like p53 or Rb. While these cells are extremely robust, easy to culture, and divide indefinitely, their highly mutated, aneuploid genomes mean they no longer perfectly represent normal, healthy human tissue.
Module 2: Membrane Biophysics
2.1 The Container of Life & Compartmentalization
Protecting order from the inevitable decay of entropy.
The cell is essentially a highly orchestrated, self-reproducing system of biochemical reactions held inside a physical container. That indispensable container is the Plasma Membrane. Without a physical boundary, the localized concentration of vital substrates, enzymes, and ATP would instantly diffuse away, increasing system entropy and culminating in immediate cellular death.

Figure 2.1: The Fluid Mosaic Model. Note the integral transmembrane proteins, cholesterol molecules, and the carbohydrate glycocalyx on the extracellular face.
2.2 Amphipathic Nature & The Hydrophobic Effect
Resolving thermodynamic conflict in an aqueous universe.
Cellular life evolved in, is filled with, and is completely surrounded by water. Consequently, all membrane architecture is dictated entirely by how lipid molecules behave when immersed in an aqueous solvent. All structural membrane lipids are fundamentally Amphipathic (from the Greek amphi = both, pathos = feeling): they contain a strongly hydrophilic (water-loving) polar head and a strongly hydrophobic (water-fearing) non-polar hydrocarbon tail.
Phosphatidylcholine
The polar head groups contain charged phosphates and amines. They readily form energetically highly favorable hydrogen bonds and electrostatic dipole-dipole interactions with the partial charges of surrounding water molecules, dissolving easily.
The uncharged, non-polar hydrocarbon tails absolutely cannot form hydrogen bonds. When forced into water, they disrupt the dynamic hydrogen-bonding network of bulk water. To compensate and maximize hydrogen bonding with themselves, the water molecules are forced to reorganize into a highly ordered, rigid, ice-like cage (a clathrate) around the lipid tail. Thermodynamics dictates that creating this artificial order causes a massive decrease in system entropy ($\Delta S < 0$), requiring a severe, unsustainable input of free energy ($\Delta G > 0$).
The Geometric Resolution: Self-Sealing Spheres
To completely avoid the massive thermodynamic penalty of the hydrophobic effect, amphipathic lipids spontaneously self-assemble into large aggregates that sequester their tails away from water, thereby maximizing the entropy of the surrounding solvent.
Because a flat, planar lipid bilayer sheet leaves its highly hydrophobic edges exposed to water along its entire perimeter, it remains energetically unstable. The only geometric solution to eliminate all exposed edges is for the planar sheet to spontaneously bend and seal upon itself, forming a continuous, closed 3D spherical compartment (a liposome). This inescapable physical imperative of thermodynamics is the primary reason cells exist as isolated, spherical compartments. Any tear in a cell membrane will spontaneously and rapidly reseal for this exact same thermodynamic reason.
2.3 Membrane Fluidity & Lipid Rafts
Phase transitions and two-dimensional liquid crystals.
The Fluid Mosaic Model (proposed by Singer and Nicolson, 1972) posits that the biological membrane is not a static, rigid solid, but a highly dynamic two-dimensional liquid crystal. The lipids and proteins are trapped securely within the bilayer plane by the hydrophobic effect, but because they are not covalently bonded to each other, they are free to diffuse laterally at high speeds.
Types of Lipid Motion
-
Frequent
Lateral Diffusion: Lipids rapidly and constantly trade places with adjacent neighbors within the same monolayer ($\sim 10^7$ times/sec).
-
Frequent
Rotation & Flexion: Spinning rapidly along their longitudinal axis (up to $500 \text{ rev/sec}$) and continuous rapid wiggling of the hydrocarbon tails.
-
Very Rare
Transverse Diffusion (Flip-Flop): Tumbling from the outer monolayer to the inner monolayer. This is energetically prohibited because it requires dragging the heavily hydrated, polar head group through the oily hydrophobic core. It occurs spontaneously less than once a month.
Modifiers of the Phase Transition ($T_m$)
Cells actively adjust their lipid composition to maintain constant fluidity despite environmental temperature changes (Homeoviscous Adaptation).
-
Hydrocarbon Tail Length: Shorter tails possess less surface area for attractive Van der Waals interactions with neighboring lipids. Less interaction equals a more fluid membrane.
-
Saturation (Double Bonds): Saturated tails (straight) pack tightly into a solid, viscous gel. Unsaturated tails contain cis-double bonds that create rigid ~30-degree kinks. These kinks act as physical spacers, preventing tight packing and keeping the membrane highly fluid.
-
Cholesterol (Animal cells only): A rigid, planar steroid ring. It acts as a paradoxical bidirectional thermal buffer. At high temperatures, its rigid structure immobilizes the adjacent lipid tails, preventing the membrane from melting or leaking. At low temperatures, its bulky shape wedges between tails, preventing them from freezing into a solid crystal.
Experimental Proof: FRAP
To conclusively prove the membrane is fluid and measure the lateral diffusion coefficient ($D$), biologists employ FRAP (Fluorescence Recovery After Photobleaching). Membrane proteins or lipids are labeled with a fluorescent tag (like GFP). A highly focused, intense laser beam is used to irreversibly bleach a small square area of the membrane on a living cell, creating a dark "hole in the fluorescence.
Under a confocal microscope, over a period of minutes, fluorescently tagged, unbleached molecules from the surrounding membrane naturally diffuse laterally into the bleached area. The recovery of fluorescence in that square proves lateral mobility, and the mathematical curve of recovery over time allows calculation of the diffusion rate.
2.4 Biogenesis & 2.5 Transverse Asymmetry
Creating the barrier and establishing the topology.
Biological membranes are never synthesized de novo (from scratch in the cytosol). They always arise from the expansion of pre-existing membranes, specifically beginning in the Endoplasmic Reticulum (ER), undergoing refinement in the Golgi apparatus, and eventually fusing with the plasma membrane.
The Biosynthetic Assembly Line
Phospholipid synthesis enzymes are permanently embedded in the ER membrane with their active sites facing the cytosol (because their substrates, fatty acids, and ATP are cytosolic). Consequently, newly synthesized lipids are inserted exclusively into the cytosolic leaflet of the ER bilayer.
Transport vesicles carry the perfectly symmetric membrane from the ER to the Golgi. The Golgi contains a different class of highly specialized, ATP-dependent enzymes called Flippases (P4-ATPases).
Unlike Scramblases, Flippases are highly specific. They actively consume ATP to forcefully extract specific aminophospholipids—namely Phosphatidylserine (PS) and Phosphatidylethanolamine (PE)—from the lumenal (outer) leaflet and flip them strictly to the cytosolic leaflet against their concentration gradient. This establishes absolute membrane asymmetry.
Asymmetric vesicles bud from the Trans-Golgi Network and undergo exocytosis, fusing with the plasma membrane. Due to the inviolable laws of topology during membrane fusion, the cytosolic face of the vesicle always remains facing the cytosol, while the lumenal face is exposed to the extracellular fluid.
Clinical Pathology: The Apoptotic "Eat Me" Signal
Because of the strict, continuous action of Flippase, the negatively charged lipid Phosphatidylserine (PS) is entirely restricted to the inner cytosolic leaflet in normal, healthy cells.
During Apoptosis (programmed cell death), intracellular executioner Caspase enzymes are activated. These caspases perform two critical, membrane-altering cuts:
1. They permanently cleave and disable Flippase.
2. They cleave and hyper-activate a plasma membrane Scramblase (TMEM16F).
With Flippase dead and Scramblase highly active, PS rapidly randomizes and becomes massively exposed on the outer extracellular leaflet. Circulating Macrophages possess specific receptors that recognize exposed PS as an unequivocal "Eat Me signal. This leads to the swift, silent phagocytosis of the dying cell without spilling its contents or triggering a damaging inflammatory immune response. In the laboratory, this PS exposure is the gold standard for detecting early apoptosis and is commonly assayed using fluorescently tagged Annexin V.
2.6 Membrane Proteins & Structural Topologies
The functional workforce embedded in the lipid sea.
If the lipid bilayer serves as the universal, impermeable factory wall, the membrane proteins function as the highly specialized doors, windows, communication antennas, and assembly line workers embedded directly within that wall. While proteins account for roughly 50% of the mass of a typical animal plasma membrane, because single protein complexes are enormously massive (often >100,000 Daltons) compared to tiny lipid molecules (~800 Daltons), there are approximately 50 lipid molecules for every 1 protein molecule.
Association Topologies
Physically inserted into the hydrophobic core of the bilayer. They absolutely cannot be extracted without completely destroying the lipid structure using harsh detergents (like SDS or Triton X-100) which solubilize the lipids into micelles.
- Transmembrane (Single-pass): Usually an $\alpha$-helix composed of 20-25 hydrophobic amino acids. The polar peptide backbone forms hydrogen bonds with itself inside the helix, hiding from the lipid tails. (e.g., Glycophorin in RBCs, Receptor Tyrosine Kinases).
- Transmembrane (Multi-pass): Multiple amphipathic $\alpha$-helices form a ring. Hydrophobic side chains face outward to the lipids, hydrophilic side chains face inward to create an aqueous pore. (e.g., Ion channels, GPCRs).
- Beta-Barrel: A $\beta$-sheet rolled into a rigid cylinder. Found extensively as Porins in the outer membranes of mitochondria and Gram-negative bacteria.
- Monolayer-Associated: Anchored horizontally to the cytosolic half via an amphipathic $\alpha$-helix.
- Lipid-Linked: The entire protein resides in the aqueous environment but is covalently bonded to a lipid anchor (like a Farnesyl group or a GPI-anchor) embedded deep in the membrane core.
These proteins do not insert into the hydrophobic core at all. They are entirely dependent on relatively weak, non-covalent electrostatic interactions and hydrogen bonds, binding to the hydrophilic domains of integral transmembrane proteins or directly to the polar lipid head groups.
Because they do not penetrate the bilayer, peripheral proteins can be gently extracted leaving the lipid membrane completely intact by simply washing the cells with high-salt solutions (e.g., 1M NaCl) or extreme pH buffers, which disrupt the electrostatic ionic bonds holding them in place.
Bioinformatics: Hydropathy Plots
Before a protein's exact 3D structure is resolved by rigorous X-ray crystallography or Cryo-EM, scientists can computationally predict if it is a transmembrane protein by analyzing its primary amino acid sequence. A Hydropathy Plot assigns a hydrophobicity index (the free energy required to transfer the amino acid side chain to water) to each amino acid.
A continuous peak of high hydrophobicity stretching for approximately 20-30 consecutive amino acids indicates a region long enough to span the 5nm lipid bilayer as an $\alpha$-helix. A complex multi-pass protein like Bacteriorhodopsin will clearly display 7 distinct, tall hydrophobic peaks on the graph, correctly predicting its 7-transmembrane topology.
2.7 The Glycocalyx & Cell Adhesion Molecules
The Sugar Coat: Protection, Lubrication & Cellular Identification.
Animal cells are not bare lipid surfaces. The vast majority of plasma membrane proteins (Glycoproteins, Proteoglycans) and roughly 10% of outer leaflet lipids (Glycolipids) have short, complex, highly branched chains of sugars (oligosaccharides) covalently attached. Because glycosylation enzymes reside exclusively inside the lumen of the ER and Golgi, this extensive, highly hydrated sugar coating—the Glycocalyx—is located strictly and exclusively on the non-cytosolic (extracellular) face of the plasma membrane.
The dense thicket of sugars provides a physical buffer zone, shielding the delicate underlying lipid bilayer from severe sheer stress, proteases, and harsh chemical environments (e.g., gastric acid).
Oligosaccharides, particularly those terminating in negatively charged Sialic Acid, are intensely hydrophilic. They absorb massive amounts of water, creating a slimy, lubricated surface crucial for preventing erythrocytes (RBCs) from adhering to capillary walls.
The immense stereochemical diversity of branched sugars creates an infinite variety of 3D structures, acting as a highly specific molecular "fingerprint critical for immune self-recognition (ABO blood groups) and sperm-egg binding.
Clinical Pathology: Leukocyte Extravasation (Neutrophil Rolling)
The glycocalyx is the primary mediator of the initial inflammatory response. During a localized bacterial infection, resident macrophages release inflammatory cytokines (e.g., IL-1, TNF-$\alpha$). These cytokines rapidly activate the endothelial cells lining nearby venules, causing them to immediately synthesize and translocate specialized transmembrane adhesion proteins called Selectins to their luminal surface.
Selectins are carbohydrate-binding proteins (lectins). Fast-moving neutrophils racing through the bloodstream possess specific matching oligosaccharide ligands (e.g., Sialyl Lewis X) highly enriched in their glycocalyx. As the neutrophil passes the infection site, its sugars snag on the newly exposed endothelial selectins like weak molecular velcro. The tremendous physical force of the blood flow constantly breaks and reforms these weak non-covalent bonds, causing the cell to drastically slow down and literally "roll along the vessel wall.
This rolling is the mandatory first step. It slows the neutrophil enough to allow subsequent, much stronger protein-protein interactions (mediated by Integrins binding to ICAMs) to firmly arrest the cell, finally enabling it to squeeze between endothelial cells (Diapedesis) and enter the infected tissue to neutralize the pathogen.
Module 3: Transport & Neurobiology
3.1 Fick's Law & The Permeability Hierarchy
The thermodynamic barriers to entry.
The hydrophobic core of the lipid bilayer acts as a ruthless thermodynamic barrier. The absolute rate at which any uncharged molecule crosses this barrier via Simple Passive Diffusion is formally described by Fick's First Law of Diffusion ($J = -D \frac{dc}{dx}$). However, biologically, this permeability coefficient is practically governed by two critical molecular properties: Size (smaller diffuses faster) and Polarity (measured by the Oil/Water Partition Coefficient, where higher lipid solubility yields exponentially faster diffusion).
| Molecule Class | Classic Examples | Permeability Status & Biological Context |
|---|---|---|
| Small Nonpolar | $O_2, CO_2, N_2$, NO, Steroid Hormones | Highly Permeable (Simple Diffusion). Dissolve readily in the hydrophobic core and pass instantly down their gradient. Critical for rapid pulmonary gas exchange and direct nuclear receptor binding by steroids (Testosterone, Cortisol) without requiring any surface receptors. |
| Small Uncharged Polar | $H_2O$, Ethanol, Glycerol, Urea | Moderately Permeable. Despite being polar, their tiny size allows them to occasionally slip between the transient gaps created by flexing lipid tails. However, this is too slow for cells requiring massive water flux (e.g., Kidney collecting ducts), which strictly require dedicated Aquaporin channels. |
| Large Uncharged Polar | Glucose, Sucrose, Amino Acids | Extremely Low Permeability. Their physical bulk prevents slipping through lipid gaps, and their polarity prevents dissolving in the core. They absolutely require specific Carrier Transporters (like GLUT1) to cross the membrane at physiologically relevant rates. |
| Ions (Highly Charged) | $H^+, Na^+, K^+, Ca^{2+}, Cl^-$ | Strictly Impermeable ($P \approx 0$). Ions exert immense electrostatic pull, permanently attracting a rigid, highly structured sphere of water molecules (Hydration Shell). To pass the non-polar core, the ion must strip off this water. The energy cost of this dehydration (Activation Energy barrier) is insurmountable by thermal energy alone. They depend 100% on protein Channels or Pumps. |
3.2 Transporters vs. Channels: Kinetic Differences
Mechanisms of membrane proteins.
To transport impermeable molecules (classes 3 and 4), cells dedicate up to 30% of their genome to encoding specialized transmembrane proteins. These transport proteins fall into two fundamentally distinct, non-overlapping mechanistic categories with vastly different kinetic profiles.
Transporters (Carriers)
The "Turnstile" Model-
Mechanism: Do not form an open pore. They must physically bind the specific solute on one side of the membrane, undergo a massive, reversible, global conformational change, and subsequently release the solute on the opposite side.
-
Specificity: Extremely high. They act like enzymes interacting with substrates. They can distinguish between enantiomers (e.g., transports biological D-Glucose perfectly but completely ignores synthetic L-Glucose).
-
Kinetics: Because they rely on physical binding and slow conformational changes, their speed is limited ($10^2 - 10^4$ molecules/sec). Crucially, they follow Michaelis-Menten kinetics and exhibit saturation. Once all "turnstiles are occupied, transport rate hits a strict maximum velocity ($V_{max}$).
-
Capability: The only class capable of performing Active Transport (pumping uphill against a gradient using ATP), as well as Passive Facilitated Diffusion.
Ion Channels
The "Trapdoor" Tunnel-
Mechanism: Form a continuous, narrow, hydrophilic pore directly through the lipid bilayer. They do not bind the solute tightly; when the "door is open, ions simply flood through in single file.
-
Specificity: High, but based strictly on physical pore diameter and the electrostatic charge at the narrowest bottleneck (the Selectivity Filter).
-
Kinetics: Extremely fast, approaching the limits of free diffusion ($10^7 - 10^8$ ions/sec)—over 10,000 times faster than the fastest transporter. They exhibit linear kinetics and virtually never reach saturation at physiological ion concentrations.
-
Capability: Can ONLY mediate Passive Transport. Ions flow strictly downhill according to their electrochemical gradient. Channels cannot "pump".
3.3 Primary & Secondary Active Transport
Fighting entropy to create cellular batteries.
A cell in perfect thermodynamic equilibrium with its environment is dead. Sustaining life requires the continuous, massive expenditure of metabolic energy to fight entropy and maintain steep chemical and electrical gradients across the plasma membrane. These gradients act as charged biological batteries, storing potential energy used to drive crucial processes like nutrient uptake, osmotic balance, and nerve impulses.
Classes of Active Pumps
Hydrolyze ATP and use the terminal phosphate to covalently phosphorylate themselves, driving a massive conformational change. (e.g., Na+/K+ pump, SERCA Ca2+ pump).
Turbine-like machines that hydrolyze ATP to pump protons (H+) into organelles, severely acidifying them (e.g., Lysosomes pH 5.0, plant vacuoles).
V-type working in reverse. Uses a steep H+ gradient flowing downhill to mechanically spin a turbine, synthesizing ATP from ADP + Pi (Inner Mitochondrial Membrane).
ATP-Binding Cassettes. Contains 2 ATP binding domains. Largest family. Pumps out toxins/drugs. Overexpression causes Multi-Drug Resistance (MDR) in cancer.
The Universal Engine: Na+/K+ ATPase Mechanism
This single ubiquitous P-type pump utilizes up to 30% of a typical animal cell's total ATP production. It is an electrogenic antiport (it pumps 3 positive charges out for every 2 it brings in, causing a net loss of 1 positive charge per cycle, directly contributing ~10% to the negative resting membrane potential). It operates via an alternating access "Ping-Pong cycle:
- E1 State (Open to Inside): The pump has a high affinity for Sodium. 3 Na+ ions bind tightly from the low-concentration cytosol into deep central pockets.
- Phosphorylation: The binding of 3 Na+ stimulates intrinsic ATPase activity. ATP is hydrolyzed to ADP, and the terminal phosphate is covalently attached to a highly conserved Aspartate residue (Asp376) on the pump itself.
- Conformational Flip (E1 $\rightarrow$ E2): The addition of the bulky, highly charged phosphate group is energetically unfavorable in the current shape. It acts as a loaded spring, forcing a massive, irreversible shape change. The pump opens to the extracellular space. Crucially, in this new shape, its affinity for Na+ plummets, causing the 3 Na+ to be forcefully ejected outward against their gradient.
- E2 State (Open to Outside): In this new E2 conformation, the pump's binding pockets are reshaped to have a high affinity for Potassium. 2 K+ ions bind from the extracellular fluid.
- Dephosphorylation & Reset: K+ binding triggers the hydrolysis and release of the covalent phosphate group. Without the phosphate forcing the E2 shape, the pump relaxes and snaps back to its original, thermodynamically stable E1 shape. This flip ejects the 2 K+ inside the cell, readying the pump for a new cycle.
Secondary Active Transport
Does not hydrolyze ATP directly. Instead, it harnesses the kinetic energy of one solute moving downhill (usually Na+, capitalizing on the massive battery created by the Na+/K+ pump) to physically drag a second solute uphill against its gradient.
Found in the apical membrane of intestinal epithelia. The immense inward pressure of Na+ forces the symporter to flip, pulling Glucose from the gut lumen into the cell against a massive concentration gradient, ensuring absolutely no nutrients are lost in feces.
Clinical: Cardiac Glycosides
Drugs such as Digitalis and Ouabain (extracted from the Foxglove plant) have been used for centuries to treat congestive heart failure. They act as deadly competitive inhibitors, binding irreversibly to the extracellular K+ binding site of the Na+/K+ pump, permanently paralyzing the engine in the E2 state.
The Pathological Cascade:- Pump stops $\rightarrow$ Intracellular [Na+] slowly rises.
- The secondary active Na+/Ca2+ Exchanger (NCX) loses its Na+ driving force and stops pumping Ca2+ out.
- Intracellular [Ca2+] builds up dangerously.
- Excess Ca2+ is sequestered in the Sarcoplasmic Reticulum.
- Upon the next action potential, massively increased Ca2+ is released, causing a much stronger, more forceful muscle contraction (Positive Inotropy), temporarily aiding a failing heart.
3.4 Ion Channels: Atomic Mimicry & Gating
Structural Biophysics and Selectivity.
Structural Biophysics: The Selectivity Filter Paradox
The Physical Impossibility: A Potassium ($K^+$) channel is highly specific for $K^+$. However, the Sodium ion ($Na^+$) is physically smaller (atomic radius 0.95 Å) than $K^+$ (1.33 Å). A physical pore large enough to allow a basketball ($K^+$) to pass through should theoretically offer no resistance to a tennis ball ($Na^+$). Yet, the channel rejects $Na^+$ with a staggering specificity of 10,000 to 1. How does biology defy macroscopic sizing logic?
KcsA Channel Atomic Mechanism
The Solution (Atomic Mimicry): In the vestibule, both ions are surrounded by a tight hydration shell of water. To pass through the extremely narrow selectivity filter, they must be completely stripped of this water, which requires a massive input of desolvation energy.
The walls of the K+ channel filter are lined with the highly electronegative Carbonyl Oxygen atoms ($C=O$) of the protein's polypeptide backbone. Evolution has rigidly fixed these atoms in a precise 3D geometric matrix that exactly mimics the specific spacing of the water molecules in a $K^+$ hydration shell.
As $K^+$ enters, it sequentially trades the oxygens of water for the oxygens of the channel wall in a perfectly seamless transition. The energetic cost of dehydration is perfectly balanced by the new bonds formed with the channel ($\Delta G \approx 0$). $K^+$ slides through effortlessly.
Because $Na^+$ is smaller, it physically cannot reach out and touch all four channel oxygens on all sides simultaneously. The energy gained from interacting with only 1 or 2 channel oxygens is woefully insufficient to pay the massive energetic penalty of stripping its original water shell. Therefore, $Na^+$ remains energetically trapped outside, rendered functionally "too fat" (due to its inseparable water shell) to fit through the hole. (Roderick MacKinnon, Nobel Prize in Chemistry, 2003).
Gating Kinetics: Controlling the Flow
Unlike leak channels, most ion channels switch between discrete conformational states (Closed $\rightarrow$ Open $\rightarrow$ Inactivated) in response to highly specific stimuli. They act as the biological equivalent of logic gates in a computer processor.
Detect changes in the electrical field. Specialized S4 transmembrane helices contain positively charged Arginine/Lysine residues spaced every third amino acid, acting as a highly sensitive "voltage sensor". When the membrane depolarizes (inside becomes more positive), electrostatic repulsion forces these helices to slide outward toward the extracellular space, mechanically pulling levers that crank the central pore open.
Open exclusively upon the physical binding of a specific chemical molecule (ligand) to an allosteric receptor site on the channel protein, forcing a conformational shape change.
Extracellular Ligands: Neurotransmitters like Glutamate, Acetylcholine, or GABA at synapses.
Intracellular Ligands: Second messengers like cAMP, cGMP (in photoreceptors), or Calcium ions generated by internal signaling cascades.
Open directly in response to physical deformation, stretching, or tension applied to the membrane. The channel is physically tethered to the internal cytoskeleton or external matrix. Mechanical force literally pulls the trapdoor open.
Examples: Stereocilia in the inner ear converting sound wave vibrations into electrical signals; Piezo channels in the skin responsible for the sensation of light touch.
3.5 Membrane Potential & The Nernst Equation
The mathematics of the Electrochemical Gradient.
For uncharged molecules, passive transport direction is dictated solely by the concentration gradient. However, for charged molecules (ions), transport is determined by the Electrochemical Gradient, which is the net sum of two distinct physical forces.
- Chemical Force (Concentration Gradient): The entropic drive pushing molecules from an area of high concentration to an area of low concentration.
- Electrical Force (Membrane Potential): The electrostatic pull. Because the inside of a resting cell is heavily negative ($\sim -60 \text{ mV}$), it exerts a massive attractive force pulling cations ($+$) inward, and a repulsive force pushing anions ($-$) outward.
Thermodynamic Equilibrium: The Nernst Equation
If a membrane is permeable to only one specific ion, that ion will flow down its concentration gradient until the electrical charge that builds up exactly opposes further chemical movement. This state of zero net thermodynamic flow is the Equilibrium Potential ($E_{ion}$), calculated by the Nernst equation:
- $R$: Universal Gas Constant (8.314 J/mol·K)
- $T$: Absolute Temperature (Kelvin)
- $z$: Valence of the ion (e.g., +1 for Na+, +2 for Ca2+)
- $F$: Faraday's Constant (96,485 C/mol)
Concentration: High Outside (145mM), Low Inside (10mM) $\rightarrow$ Pushes IN.
Electrical: Inside is negative (-60mV) $\rightarrow$ Pulls IN.
Result: Massive Inward Force
Both forces align perfectly. If a Na+ channel opens, Na+ violently rushes into the cell (driving explosive Depolarization). The Nernst potential is $E_{Na} \approx +60 \text{ mV}$.
Concentration: High Inside (140mM), Low Outside (5mM) $\rightarrow$ Pushes OUT.
Electrical: Inside is negative (-60mV) $\rightarrow$ Pulls IN.
Result: Weak Outward Force
The forces actively fight each other. The chemical outward push is slightly stronger than the electrical inward pull. Net result is a slow, weak outward leak. The Nernst potential is $E_{K} \approx -90 \text{ mV}$.
3.6 Neurobiology: Action Potentials & Synaptic Transmission
The Hodgkin-Huxley Model and SNARE Exocytosis.
The nervous system exploits these established ionic gradients to transmit information rapidly over vast distances (up to a meter down a human leg) via electrical impulses called Action Potentials, followed by chemical transmission across synapses.
The Action Potential (Hodgkin-Huxley Model)
An action potential is an all-or-nothing, self-propagating wave of massive depolarization that travels down an axon without any loss of amplitude, driven entirely by the highly orchestrated, sequential opening and closing of Voltage-Gated Ion Channels.
-
1. Resting State (-60mV)
Established primarily by the continuous activity of the Na+/K+ pump and the presence of constitutively open K+ Leak Channels. K+ slowly leaks out, leaving impermeable negative anions (proteins, nucleic acids) behind, creating a stable negative interior.
-
2. Depolarization (The Spike)
A stimulus (e.g., neurotransmitter binding at dendrites) causes a slight initial depolarization. If this reaches the critical Threshold (-40mV), the S4 voltage sensors in Voltage-Gated Na+ Channels pop open instantly. Na+ violently rushes IN driven by its massive electrochemical gradient. This further depolarizes the membrane, opening even more Na+ channels in a rapid positive feedback loop. The membrane potential reverses polarity, shooting up to +40mV.
-
3. Inactivation & Absolute Refractory Period
Crucially, at the peak (+40mV), Na+ channels do not simply close. An intrinsic globular protein domain swings in and physically plugs the open pore from the cytosolic side (Ball-and-chain inactivation). The channel is locked and dead to further stimulation. It cannot reopen until the membrane fully repolarizes. This forces the action potential to propagate in only one direction (forward) because the membrane behind it is completely refractory.
-
4. Repolarization (Falling Phase)
Slightly slower, Voltage-Gated K+ Channels open. K+ rapidly rushes OUT of the cell down its gradient, carrying positive charge away and quickly restoring the negative resting potential.
The Chemical Synapse & SNARE Exocytosis
Electrical signals absolutely cannot jump the physical 20nm gap (synaptic cleft) between neurons. The electrical signal must be temporarily converted into a chemical messenger (neurotransmitter) via highly regulated vesicular exocytosis, and then converted back into an electrical signal on the other side.
- 1. Arrival & Calcium Influx: The Action Potential reaches the presynaptic axon terminal. The sudden depolarization opens Voltage-Gated Calcium ($Ca^{2+}$) Channels. Because extracellular Ca2+ is maintained at levels 10,000 times higher than intracellular levels, Ca2+ violently floods into the terminal.
- 2. The Calcium Sensor: The Ca2+ binds to a highly specialized sensor protein called Synaptotagmin located on the membrane of synaptic vesicles (which are packed with neurotransmitters).
- 3. The SNARE Complex Fusion: Synaptotagmin binding releases a block (Complexin), allowing the SNARE proteins to interact. A single v-SNARE (Synaptobrevin) on the vesicle tightly intertwines with two t-SNAREs (Syntaxin and SNAP-25) on the plasma membrane. They form an incredibly stable 4-helix bundle. Like a biological winch, this zippering mechanically forces the two lipid bilayers within 1.5nm of each other, forcibly squeezing out the hydration water molecules and triggering instantaneous membrane fusion. Neurotransmitters are dumped into the cleft.
- 4. Post-Synaptic Transduction: Neurotransmitters diffuse across the cleft in microseconds and bind to Ligand-Gated Ion Channels on the post-synaptic dendrite.
Neurotransmitters like Glutamate or Acetylcholine bind and open non-specific Cation channels. $Na^+$ rushes IN, bringing positive charge. This depolarizes the local membrane, pushing it closer to the threshold required to fire a new action potential.
Neurotransmitters like GABA or Glycine bind and open specific Anion channels. $Cl^-$ rushes IN, bringing negative charge. This hyperpolarizes the membrane (makes it more negative than resting), pulling it further away from the firing threshold, effectively silencing the neuron.
Clinical Pathology: Botulinum & Tetanus Neurotoxins
The extreme precision of the SNARE fusion machinery is actively exploited by some of the deadliest toxins known to mankind, produced by Clostridium bacteria.
Botulinum Toxin (Botox) acts as a highly specific zinc-dependent endoprotease. Once taken up into the presynaptic terminal of motor neurons, it actively cleaves specific SNARE proteins (like SNAP-25). Without intact SNAREs, synaptic vesicles containing Acetylcholine can dock but absolutely cannot fuse. The muscle never receives the signal to contract, resulting in flaccid paralysis (and cosmetically, the elimination of wrinkles).
Tetanus Toxin uses the exact same enzymatic mechanism (cleaving SNAREs), but it specifically targets inhibitory interneurons (Renshaw cells) in the spinal cord that release GABA. By preventing the release of the "brakes," the motor neurons fire uncontrollably, resulting in severe, rigid, spastic paralysis (lockjaw).
Advanced Neuroscience: Long-Term Potentiation (LTP) & Memory
How does a physical synapse "learn"? Learning and memory are believed to be physically stored as the persistent, long-lasting strengthening of specific synapses, a phenomenon called Long-Term Potentiation (LTP), primarily studied in the Hippocampus. It relies on the elegant molecular interplay of two different Glutamate receptors: AMPA and NMDA.
- Baseline Transmission: Normal glutamate release only opens AMPA Receptors ($Na^+$ enters, causing mild depolarization). The NMDA Receptor is completely blocked by a giant $Mg^{2+}$ ion physically wedged in its pore.
- Tetanic Stimulation (The Learning Event): High-frequency, intense, repetitive stimulation causes massive glutamate release and strong, prolonged depolarization via the AMPA receptors.
- The Coincidence Detector: The strong internal positive charge electrostatically repels and blows the $Mg^{2+}$ plug out of the NMDA receptor. The NMDA receptor acts as a biological AND-gate: it requires BOTH glutamate binding AND significant membrane depolarization to open.
- Calcium Influx & Plasticity: The unblocked NMDA receptor is highly permeable to $Ca^{2+}$. Calcium floods the postsynaptic spine, activating calcium-dependent kinases (CaMKII, PKC). These kinases trigger the rapid insertion of *more* AMPA receptors into the postsynaptic membrane from internal endosomes, and physically enlarge the dendritic spine (structural plasticity).
- The Result (Memory Formation): The synapse is permanently remodeled. The next time a weak, baseline signal arrives, the larger number of AMPA receptors generates a much larger EPSP. The neural pathway has been physically strengthened; a memory has been formed.
Module 4:
Organelles & Traffic
4.1 Compartmentalization
The Evolutionary Leap of Eukaryotes.
Prokaryotes generally possess only a single compartment (the cytoplasm) where all biochemical reactions proceed simultaneously. Eukaryotic cells, however, possess volumes 1,000 to 10,000 times larger. In such a massive volume, relying purely on simple thermal diffusion for an enzyme to encounter its specific substrate is thermodynamically disastrous and far too slow to sustain complex multicellular life.
Evolution solved this profound physical crisis through Compartmentalization: dividing the intracellular space into numerous closed, specialized, membrane-bound volumes called organelles. This extensive use of intracellular lipid membranes is theorized to have originated via two primary pathways: the autogenous infolding of the ancestral plasma membrane (yielding the nuclear envelope and ER network), and endosymbiosis (yielding mitochondria and chloroplasts).
The Three Crucial Thermodynamic Advantages
Specific metabolic pathways require precise sets of enzymes, substrates, and cofactors. By confining them within a tiny organelle volume, the cell dramatically increases their local concentration. According to the laws of mass action, this shifts the reaction kinetics favorably, completely overcoming the massive dilution problem of a large eukaryotic cell.
Compartments establish independent chemical parameters impossible to maintain in the bulk cytosol. For example, Lysosomes utilize V-type ATPases to actively pump protons, maintaining a harsh internal pH of ~5.0. This acidity is strictly required for the function of resident acid hydrolases, while the cytosol remains safely at pH 7.2.
Isolating highly dangerous biochemical reactions. If potent proteases, nucleases, or reactive oxygen species (like $H_2O_2$ generated continually in peroxisomes) were allowed to float freely in the cytosol, they would indiscriminately destroy the cell's own DNA and vital structural proteins.
Visualizing Compartmentalization

Figure 4.1: Animal Cell Architecture. Note the extensive dynamic membrane networks and lack of a rigid cell wall.

Figure 4.2: Plant Cell Architecture. Characterized by a rigid cellulose wall, a massive central vacuole for maintaining turgor pressure, and chloroplasts for autotrophy.
Semantic Distinction: Cytoplasm vs. Cytosol
These terms are frequently confused in literature but represent distinct physical spaces in cell biology.
- Cytoplasm: The entire contents of the cell contained within the plasma membrane, excluding the nucleus. It is a comprehensive term that includes all membrane-bound organelles, cytoskeletal filaments, and the fluid medium itself.
- Cytosol: The highly concentrated aqueous, gel-like fraction of the cytoplasm that remains after all organelles and insoluble cytoskeletal elements have been physically removed (usually via ultracentrifugation). It constitutes roughly 50% of the total cell volume and is the primary site of protein translation, glycolysis, and major intracellular signaling cascades.
4.2 Organelle Master Reference
Comprehensive classification and functions.
Eukaryotic organelles are broadly classified by their bounding structure: Double-membrane (Envelope), Single-membrane, or Membraneless (macromolecular condensates formed via liquid-liquid phase separation). Each possesses a highly specific proteome dictating its unique biochemical function.
| Organelle | Boundary | Primary Function | Detailed Mechanisms & Clinical Notes |
|---|---|---|---|
| Nucleus | Double Membrane (Envelope) | Storage of genomic DNA, control of transcription, and mRNA processing. | Inner membrane is lined by the Nuclear Lamina (Lamin A/B/C) providing structural support. Outer membrane is continuous with the Rough ER. Transport is strictly gated by massive Nuclear Pore Complexes (NPCs). |
| Rough ER | Single Membrane (Network) | Translation, folding, and initial N-linked glycosylation of secretory/membrane proteins. | Studded with 80S Ribosomes on the cytosolic face. Accumulation of misfolded proteins triggers the Unfolded Protein Response (UPR). Contains SEC61 translocon. |
| Smooth ER | Single Membrane (Tubules) | Lipid synthesis, xenobiotic detoxification, and intracellular calcium storage. | Highly abundant in Hepatocytes utilizing Cytochrome P450 for drug detoxification. In muscle cells, it is specialized as the Sarcoplasmic Reticulum for rapid $Ca^{2+}$ release via RyR receptors driving contraction. |
| Golgi App. | Single Membrane (Stacked) | Post-translational modification, sorting, and packaging of ER products. | Proteins enter the Cis face, are sequentially modified (e.g., complex glycosylation, O-linked glycosylation) in the Medial cisternae, and sorted at the Trans Golgi Network (TGN). M6P tags target to lysosomes. Follows the Cisternal Maturation Model. |
| Lysosomes | Single Membrane | Intracellular digestion and Autophagy (self-eating). | Lumen is maintained at pH 5.0 via a V-type $H^+$-ATPase. Contains ~40 acid hydrolases. Genetic enzyme deficiencies result in lethal Lysosomal Storage Diseases (e.g., Tay-Sachs, Gaucher, Niemann-Pick). |
| Endosomes | Single Membrane | Sorting stations for endocytosed material. | Early endosomes (marked by Rab5) receive cargo. They mature into Late Endosomes (marked by Rab7), becoming more acidic before fusing with lysosomes. Recycling Endosomes (Rab11) return receptors to plasma membrane. |
| Mitochondria | Double Membrane | ATP production via Oxidative Phosphorylation. | Originates from an endosymbiotic $\alpha$-proteobacterium. Contains its own circular mtDNA and 70S ribosomes. Inner membrane forms cristae to maximize surface area for the Electron Transport Chain. Releases Cytochrome C to trigger intrinsic apoptosis. |
| Peroxisomes | Single Membrane | $\beta$-oxidation of very-long-chain fatty acids (VLCFAs) and $H_2O_2$ neutralization. | Oxidative enzymes produce highly toxic $H_2O_2$, which is immediately degraded to water and oxygen by Catalase. Biogenesis failure causes Zellweger syndrome. Proteins imported via PTS1/2 signals. |
| Ribosomes | Membraneless | Translation of mRNA into polypeptide chains. |
Massive macromolecular complexes of rRNA and proteins. Eukaryotic 80S (40S + 60S). Assembled in the nucleolus. Exist free in the cytosol or bound to the RER.
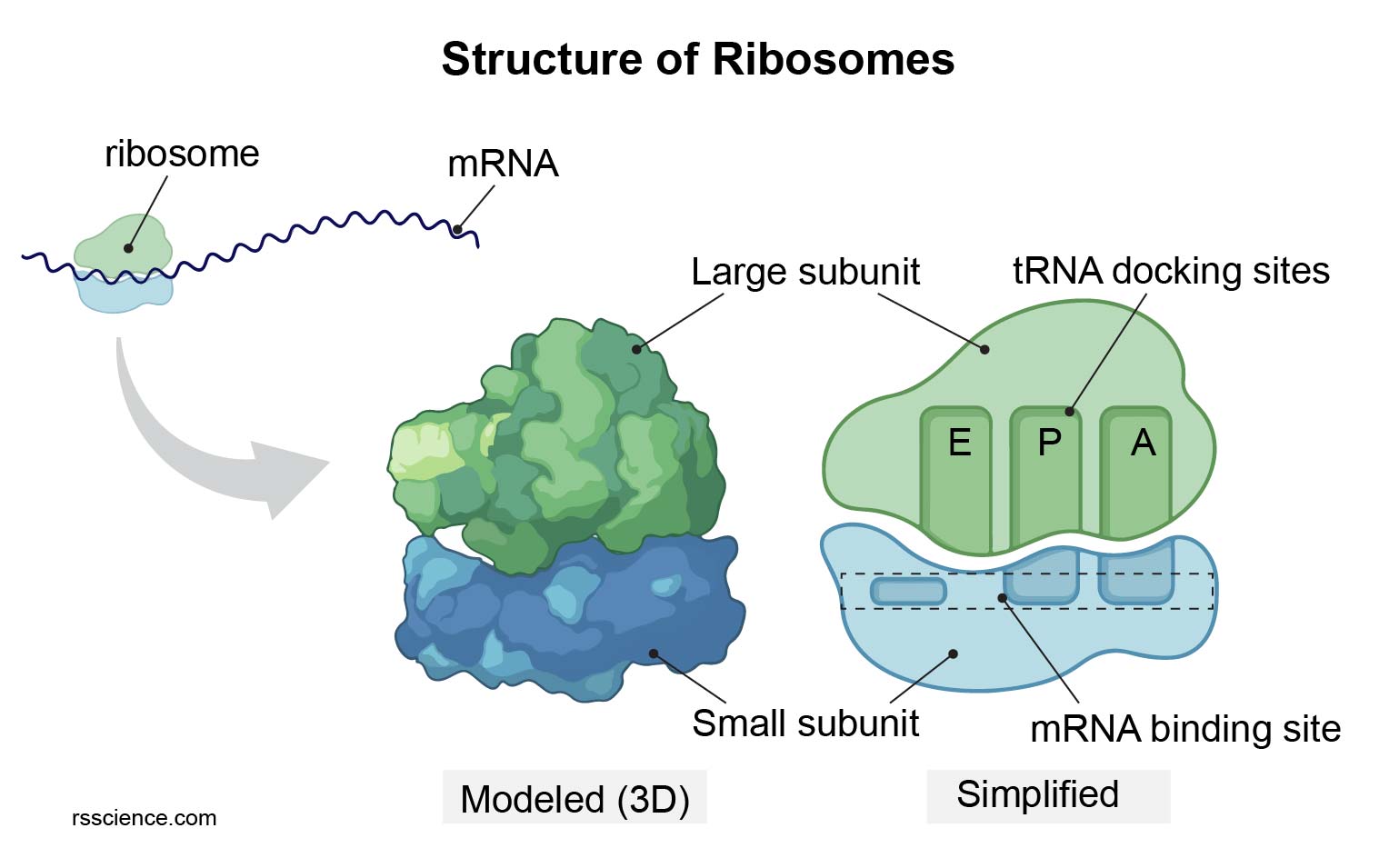
|
| Proteasome | Membraneless | Targeted degradation of misfolded or unneeded proteins. | Massive 26S barrel-shaped protease complex. Proteins must be specifically tagged with a poly-Ubiquitin chain (via E1, E2, E3 ligases) to be recognized and destroyed (Ubiquitin-Proteasome System). |
4.3 The Endomembrane System & 4.4 Protein Sorting
The Intracellular Logistics Network.
The Endomembrane System is a dynamic, structurally continuous and vesicular-interconnected network composed of the Nuclear Envelope, ER, Golgi Apparatus, Endosomes, Lysosomes, and the Plasma Membrane. Materials continually flow between these compartments via targeted vesicular transport. Crucially, Mitochondria and Chloroplasts are evolutionarily distinct endosymbionts and exist entirely outside this specific vesicular network.
How does a newly synthesized protein know its precise destination within this complex city? The answer lies in Signal Sequences (Sorting Signals)—continuous stretches of 15-60 amino acids intrinsic to the protein's primary structure that act as an unforgeable molecular "zip code".
Targeting the ER: Co-translational Translocation
Proteins destined for secretion, the plasma membrane, or lysosomes must enter the ER as their first step. Unlike mitochondrial import (which occurs post-translationally, after the protein is fully synthesized in the cytosol), import into the ER occurs co-translationally (simultaneously with translation) to prevent the highly hydrophobic membrane-spanning domains from aggregating catastrophically in the aqueous cytosol.
The Signal Recognition Particle (SRP) Cycle
-
1. Recognition & Translation Pause
As the ribosome translates mRNA in the cytosol, a highly hydrophobic sequence (the ER Signal Sequence, typically 8-15 non-polar amino acids) emerges from the ribosomal exit tunnel. A large ribonucleoprotein complex called the Signal Recognition Particle (SRP) immediately recognizes and binds to this sequence. This binding sterically blocks the elongation factor binding site on the ribosome, strictly pausing protein synthesis. This critical pause prevents the hydrophobic domains from misfolding or aggregating in the aqueous cytosol before reaching the membrane.
-
2. Docking at the ER Membrane
The entire SRP-Ribosome-mRNA-nascent chain complex diffuses to the rough ER membrane. Here, the SRP firmly binds to the transmembrane SRP Receptor. This highly specific interaction precisely aligns the ribosome directly over a closed protein pore complex called the Sec61 Translocon.
-
3. Translocation & Processing
The SRP and its receptor both hydrolyze GTP, causing SRP to release the signal sequence and dissociate, allowing translation to resume. The growing polypeptide chain is physically pushed (via the mechanical force of translation) through the aqueous pore of the open Sec61 channel into the ER lumen. An adjacent membrane enzyme, Signal Peptidase, cleaves the signal sequence. Inside the lumen, chaperone proteins (like BiP, an Hsp70 homolog, and Calreticulin) bind the chain, assisting in proper 3D folding and initiating core N-linked glycosylation.
4.5 Vesicular Logistics & Fusion
The molecular delivery trucks and SNARE mechanics.
Transport between compartments requires the formation of spherical Transport Vesicles. This process must be highly regulated: the vesicle must physically deform the flat membrane to bud off, incorporate the correct cargo, travel along microtubule highways, and definitively fuse only with the correct target membrane.
The Coat Proteins (Vesicle Budding)
Drives budding from the ER to the Golgi (Cis face). Assembly is initiated by the Sar1 GTPase. Cargo includes newly synthesized proteins bound for secretion or later compartments.
Drives budding from the Golgi back to the ER. Essential for retrieving escaped ER-resident proteins (like BiP or PDI) that carry a C-terminal KDEL retention signal.
Mediates transport from the Trans-Golgi to Lysosomes, and inward from the Plasma Membrane to Endosomes. Clathrin triskelions self-assemble into a rigid geometric cage to force membrane curvature. Requires the GTPase Dynamin to physically pinch off the vesicle neck.
Target Recognition: The SNARE Hypothesis
How does a vesicle specifically recognize its correct destination amidst the crowded cytoplasm, and how does it generate the immense physical force required to merge two strongly repelling lipid bilayers?
-
1. Tethering (Rab GTPases)
Each vesicle surface is tagged with a specific Rab GTPase (the "ID badge"). The target membrane possesses long, thread-like Tethering Proteins (Rab effectors) that specifically recognize and bind the Rab protein, acting like a molecular fishing line to catch the vesicle from the cytosol at a distance.
-
2. Docking (The SNARE Complex)
Once tethered and pulled close, a single transmembrane $\alpha$-helix on the vesicle (v-SNARE, e.g., Synaptobrevin) makes contact with a complex of three $\alpha$-helices on the target membrane (t-SNARE, e.g., Syntaxin and SNAP-25).
-
3. Fusion (The Molecular Winch)
The v-SNARE and t-SNAREs possess hydrophobic surfaces. To avoid water, they rapidly intertwine, zippering up into an incredibly stable 4-helix bundle (the trans-SNARE complex). The kinetic energy released by this violent winding acts like a mechanical winch, forcibly pulling the two lipid bilayers to within 1.5 nm of each other. This immense physical pressure literally squeezes out the hydration shells of water molecules between the lipids, triggering spontaneous, instantaneous membrane fusion.
-
4. Disassembly (NSF ATPase)
The resulting cis-SNARE complex (now entirely on the target membrane) is so tightly bound it requires an accessory hexameric AAA+ ATPase called NSF (along with $\alpha$-SNAP proteins) to forcibly pry the helices apart, consuming massive amounts of ATP, allowing the v-SNAREs to be recycled back to the donor compartment.
4.6 Endocytosis Pathways
Internalizing the extracellular environment.

Figure 4.3: Bulk transport mechanisms. Exocytosis for massive secretion; Endocytosis for internalization and membrane recycling.
Phagocytosis
"Cell Eating"
Ingestion of massive solid particles (>250nm) such as whole invading bacteria, dead cells, or cellular debris. This process is highly regulated, receptor-triggered, and strictly restricted to specialized immune professional cells (Macrophages, Neutrophils, Dendritic cells). It is driven by large-scale, actin-mediated membrane extensions (pseudopods) that engulf the target, forming a massive Phagosome which fuses with lysosomes for destruction.
Pinocytosis
"Cell Drinking"
The continuous, non-specific, constitutive uptake of extracellular fluid and dissolved solutes via small vesicles (<150nm). It occurs constantly in virtually all eukaryotic cells. It is crucial not only for nutrient uptake but for balancing the massive amount of lipid membrane added to the cell surface during continuous exocytosis (membrane recycling).
Receptor-Mediated
Highly Selective
Specific macromolecules bind to complementary transmembrane receptors, which then rapidly cluster via adaptins into Clathrin-coated pits before invaginating. This highly efficient mechanism concentrates rare extracellular cargo up to 1000-fold compared to simple fluid-phase pinocytosis.
Clinical Mechanism: LDL Cholesterol Uptake
Because cholesterol is an entirely hydrophobic lipid, it travels through the aqueous human bloodstream packaged inside massive Low-Density Lipoprotein (LDL) particles. Cells requiring cholesterol for membrane synthesis import these particles strictly via Receptor-Mediated Endocytosis.
- LDL particles bind tightly to specific LDL Receptors exposed on the extracellular plasma membrane.
- Adaptin proteins (AP2) bind the cytosolic tail of the receptor, recruiting Clathrin triskelions to assemble a polyhedral cage, forcing the membrane to invaginate and bud off (pinched by Dynamin) as a coated vesicle.
- The Clathrin coat is rapidly shed via uncoating ATPases. The naked vesicle fuses with an Early Endosome.
- The interior of the endosome is actively acidified (pH ~6.0) by V-type ATPases. This mild acidity induces a massive structural conformational change in the LDL receptor, forcing it to release the LDL particle.
- The empty receptors bud off in a small recycling vesicle and return to the plasma membrane to be used again (up to 100 trips).
- The released LDL particle is delivered to the Lysosome (pH 5.0), where potent acid hydrolases degrade the ApoB protein coat and release free cholesterol into the cytosol for cellular use.
A severe autosomal dominant genetic disorder caused by mutations in the LDL receptor gene (often creating a defective binding domain, or a defective cytosolic tail that cannot recruit AP2/Clathrin). Cells cannot internalize LDL from the blood, causing cholesterol to accumulate to massive, toxic levels in circulation, leading to severe atherosclerosis, xanthomas, and fatal myocardial infarctions early in life.
4.7 Energy & Degradation
Mitochondria and Peroxisomes.
Mitochondria
The PowerhouseProduces the vast majority of the cell's ATP via Cellular Respiration (Oxidative Phosphorylation). According to the Endosymbiotic Theory, mitochondria originated from an aerobic $\alpha$-proteobacterium engulfed by an ancestral eukaryote. Evidence includes their double membrane, possession of their own circular DNA (mtDNA), and prokaryote-like 70S ribosomes.


Figure 4.4: Mitochondrial ultrastructure and metabolic integration. Note the highly folded cristae maximizing surface area for the ETC.
Highly impermeable (rich in the unique lipid Cardiolipin). Extensively folded into Cristae to maximize the surface area available for the multi-protein Electron Transport Chain (ETC) complexes and the ATP Synthase rotor machinery.
The dense internal aqueous compartment. Contains enzymes for the Citric Acid (Krebs) Cycle and $\beta$-oxidation of fatty acids, alongside multiple copies of the mtDNA genome and ribosomes.
Mitochondrial Dynamics & Mitophagy
Mitochondria are highly plastic, constantly merging and splitting in a delicate balance to adapt to cellular energy demands and facilitate strict quality control.
Peroxisomes

Small, single-membrane vesicles containing intense oxidative enzymes. They execute the $\beta$-oxidation of Very-Long-Chain Fatty Acids (VLCFAs) down to manageable sizes before sending them to mitochondria for further processing.
The oxidation reactions generate massive amounts of Hydrogen Peroxide ($H_2O_2$), an extremely toxic reactive oxygen species. Peroxisomes contain large crystalline cores of the enzyme Catalase, which instantly neutralizes it into harmless water and oxygen ($2H_2O_2 \rightarrow 2H_2O + O_2$).
A fatal getic disorder caused by mutations in PEX genes required for peroxisome biogenesis. Without functional peroxisomes, VLCFAs accumulate to toxic levels in the blood and tissues, causing devastating neurological demyelination and infant mortality.
Chloroplasts

Found exclusively in plant cells and specific protists. Like mitochondria, they possess a double membrane, circular DNA, and originated from an endosymbiotic cyanobacterium. They execute Photosynthesis, utilizing the green pigment chlorophyll embedded in internal membrane stacks (Thylakoids, forming Grana) to harvest photon energy and fix atmospheric $CO_2$ into sugars within the fluid Stroma via the Calvin Cycle.
Module 5:
The Nucleus
5.1 The Command Center & 5.2 Morphology
Safeguarding the code of life.
The nucleus is the definitive structural hallmark of eukaryotic life. By physically sequestering the fragile genomic DNA within a robust double-membrane fortress, the cell achieves precise spatial and temporal control over gene expression. This profound architectural division decisively uncouples the processes of transcription (synthesizing mRNA from DNA) from translation (synthesizing proteins at ribosomes), enabling complex RNA splicing and epigenetic processing mechanisms impossible in prokaryotes.
Genetic Storage
Contains nearly the entirety of the cell's genetic blueprint, organized into linear chromosomes. If stretched end-to-end, the DNA from a single human nucleus would measure approximately 2 meters in length, yet it is intricately packaged via histones into a highly ordered sphere only ~10 µm in diameter.
Nuclear Variations
Most cells are mononucleated. However, skeletal muscle fibers and bone-resorbing osteoclasts are Syncytia (multinucleated via cell fusion). Megakaryocytes are Coenocytes (multinucleated via nuclear division without cytokinesis). Mammalian red blood cells are entirely Anucleate, having ejected their nucleus during maturation to maximize space for hemoglobin.
Clinical Morphology
While typically spherical (e.g., hepatocytes), nuclei can be heavily lobulated to allow severe cellular deformation through narrow capillaries (e.g., neutrophils). In oncology pathology, an increased Nucleus-to-Cytoplasm ratio (High N:C ratio) and severe size/shape variation (Anisonucleosis) are critical, definitive diagnostic hallmarks of malignancy.
5.3 Nuclear Envelope & Nuclear Lamina
The Double Fortress.
The nucleus is bounded by the Nuclear Envelope, consisting of two concentric lipid bilayers. Although they physically merge at the periphery of the nuclear pores, their protein compositions and functional roles are entirely distinct.
Faces the internal nucleoplasm. It contains unique integral membrane proteins (e.g., LBR, Emerin) that act as crucial attachment sites for the underlying Nuclear Lamina and for silent regions of chromatin (forming Lamina-Associated Domains, or LADs).
Faces the cytoplasm and is physically continuous with the membrane of the Rough ER. Like the RER, its cytosolic surface is studded with active ribosomes synthesizing proteins directly into the perinuclear space.
Contains SUN (inner) and KASH/Nesprin (outer) domain proteins that bridge the envelope, physically coupling the nucleus directly to the cytoplasmic cytoskeleton (Actin/Microtubules) for positioning and mechanotransduction.
The Nuclear Lamina
A dense, highly organized 2D fibrillar meshwork lining the inner surface of the inner nuclear membrane. It is constructed from specialized intermediate filaments called Lamins (A, B, and C).
- Provides the nucleus with immense mechanical strength and dictates its overall spherical shape.
- Serves as a rigid scaffold to tether and organize inactive heterochromatin to the nuclear periphery.
- During cell division, massive phosphorylation of Lamins by M-Cdk causes the immediate, catastrophic disassembly of the lamina meshwork, leading to the necessary fragmentation of the nuclear envelope before mitosis can proceed.
Hutchinson-Gilford Progeria Syndrome is a devastating, accelerated aging disorder. It is caused by a dominant point mutation in the LMNA gene, creating a cryptic splice site that produces an aberrant, truncated Lamin A protein called Progerin. Because progerin cannot be properly processed (it retains its farnesyl lipid anchor), it becomes permanently embedded in the inner membrane, destroying the lamina meshwork. This results in severe nuclear blebbing, catastrophic genomic instability, loss of heterochromatin, and premature cell senescence, leading to cardiovascular death in the teens.
5.4 Nuclear Pores & 5.5 The Ran-GTP Cycle
The 120 MDa Gating Machine.
The nuclear envelope is perforated by thousands of colossal, octagonal channels known as Nuclear Pore Complexes (NPCs). Composed of approximately 30 distinct proteins called Nucleoporins (Nups) arranged in multiples of eight, a single vertebrate NPC weighs a staggering ~120 Megadaltons.
NPC Architecture
The Molecular Sieve (FG-Nups)
Small water-soluble molecules a ions ($< 40$ kDa) can freely and rapidly diffuse passively through the aqueous pore. However, the central channel is choked with completely unstructured, intrinsically disordered proteins containing repeating motifs of Phenylalanine and Glycine (FG-repeats). These hydrophobic domains aggregate to form a dense, gel-like thicket (a hydrogel) via liquid-liquid phase separation. This physical mesh absolutely prevents the passive diffusion of massive macromolecules like RNA polymerases and mRNA complexes.
Active Transport Mechanism
To pass the gate, large proteins require a specific molecular ticket: a Nuclear Localization Signal (NLS) (e.g., a short stretch of positively charged basic amino acids like Lysine and Arginine). This signal is recognized by soluble cytosolic receptor proteins called Importins. The Importin binds the cargo, and utilizes its own affinity for the FG-repeats to temporarily dissolve the hydrogel mesh, effectively "walking the massive cargo through the pore in a fully folded, 3D state.
The Directionality Engine: Ran-GTP Gradient
If Importins can diffuse randomly through the pore, what prevents them from carrying the cargo right back out? Directional transport is energetically driven by a steep spatial concentration gradient of the small monomeric GTPase protein, Ran.
Inside the nucleus, an accessory enzyme called Ran-GEF (Guanine Nucleotide Exchange Factor, or RCC1) is permanently tethered to the chromatin. It continuously strips GDP from Ran and replaces it with abundant intracellular GTP.
When the Importin-Cargo complex arrives here, the high concentration of Ran-GTP forces it to bind to the Importin. This binding triggers a severe allosteric shift in the Importin, forcing it to drop its cargo exclusively inside the nucleus.
In the cytosol, a different enzyme called Ran-GAP (GTPase Activating Protein) floats freely (or is tethered to the cytoplasmic fibrils of the NPC).
When the empty Importin (still bound to Ran-GTP) exits the pore back into the cytosol, Ran-GAP immediately triggers Ran to hydrolyze its GTP into GDP. Ran-GDP has zero affinity for Importin and falls off. The Importin is now reset, empty, and ready to bind a new cargo NLS.
5.6 Nucleolus & 5.7 Epigenetics
Ribosome Factories and The Histone Code.
The" Nucleolus
The nucleolus is a massive, highly dynamic, non-membrane-bound condensate located within the nucleoplasm. It is the exclusive site for Ribosome Biogenesis. It forms via liquid-liquid phase separation (LLPS) around specific chromosomal loops containing tandem arrays of rDNA genes, known as Nucleolar Organizer Regions (NORs), found on the short arms of human acrocentric chromosomes (13, 14, 15, 21, and 22).
The pale inner core. Contains the transcriptionally inactive rDNA loops and pre-assembled RNA Polymerase I machinery waiting for activation.
Surrounds the FC. The site of furious, active transcription of the large 45S pre-rRNA transcript and initial snoRNA-mediated processing.
The massive outer shell. Represents pre-ribosomal particles assembling with imported ribosomal proteins before nuclear export to the cytosol.
Chromatin Packaging & The Histone Code
DNA does not exist naked in the cell. It is intricately spooled around highly conserved, positively charged protein octamers (two copies each of H2A, H2B, H3, and H4) to form Nucleosomes (the "beads on a string structure). The unstructured N-terminal tails of these histones protrude outward and are subjected to vast arrays of reversible post-translational modifications.
| Modification | Enzymes | Biophysical Effect | Transcriptional Status |
|---|---|---|---|
| Acetylation (Lysine) |
HATs (Writers) | Physically neutralizes the positive charge of Lysine, weakening the electrostatic grip on the negative DNA phosphate backbone. Loosens DNA. | Euchromatin (Active) |
| Deacetylation | HDACs (Erasers) | Restores the positive charge on Lysine tails. DNA wraps tightly around the octamer, physically blocking RNA Polymerase access. | Heterochromatin (Silent) |
| Methylation (Lysine / Arginine) |
HMTs | Does not alter charge. Instead, creates a highly specific binding site for reader proteins (e.g., HP1 binds to H3K9me3 to condense chromatin). | Context Dependent (H3K9me3 = Silent, H3K4me3 = Active) |
Module 6:
The Cytoskeleton
6.1 Architectural Overview & Classes
The dynamic bones and muscles of the cell.
The cytoplasm is supported by an immensely complex, highly dynamic 3D network of protein filaments known as the Cytoskeleton. Unlike a vertebrate skeleton, it is not permanent; it constantly self-assembles and dismantles via rapid polymerization and depolymerization of small, soluble protein subunits. It dictates cell shape, resists catastrophic shear stress, drives massive intracellular logistics via motor proteins, and produces the immense physical forces required for cellular crawling and cytokinesis.
Figure 6.1: Immunofluorescence visualization highlighting the distinct spatial organization of the three major cytoskeletal networks.
The Three Cytoskeletal Pillars
| Feature | Actin Filaments (Microfilaments) | Microtubules | Intermediate Filaments |
|---|---|---|---|
| Protein Subunit | Globular Actin (G-actin) | $\alpha$-Tubulin / $\beta$-Tubulin Heterodimer | Fibrous proteins (e.g., Keratins, Vimentin, Lamins) |
| Structure & Diameter | Double-stranded flexible helix. ~7 nm (Thinnest) |
Rigid, hollow cylinder of 13 protofilaments. ~25 nm (Thickest) |
Staggered coiled-coil rope-like bundles. ~10 nm (Medium) |
| Polarity | Yes (Plus & Minus ends) | Yes ($\beta$-tubulin exposed at Plus end) | No (Antiparallel tetramer assembly) |
| Nucleotide Dependency | ATP binding and hydrolysis controls stability. | GTP binding ($\beta$-tubulin only) prevents catastrophe. | None required for assembly. |
| Mechanical Property | Semi-flexible. Easily ruptured if unsupported. Forms rigid bundles via cross-linking proteins. | Highly rigid. Best at resisting massive compression forces (like steel girders). Ruptures upon extreme stretching. | Incredible Tensile Strength. Extremely flexible. Acts like a bungee cord to resist severe tissue stretching and shear stress. |
| Primary Cellular Roles | Cell cortex shape, Lamellipodia (Crawling), Cytokinesis Contractile Ring, Muscle Contraction. | Long-range Vesicle/Organelle tracking highways, Mitotic Spindle, Cilia/Flagella beating. | Purely mechanical strength. Desmosomes (cell-to-cell). Nuclear envelope architecture. |
| Pharmacological Toxins | Phalloidin (Stabilizes), Cytochalasin (Depolymerizes). | Taxol (Stabilizes, Chemo), Colchicine (Depolymerizes). | Generally none (highly stable). |
6.2 Polymer Physics & Thermodynamics
The Math of Assembly: $C_c$ and Treadmilling.
The assembly of Actin and Microtubules is a non-covalent, fully reversible thermodynamic process. The rate at which the polymer grows (adds subunits at the end) is strictly dependent on the concentration of free monomers colliding with the filament end ($k_{on} \cdot [C]$). Conversely, the rate at which subunits fall off the end ($k_{off}$) is a constant, intrinsic structural property of the polymer end, entirely independent of concentration.
The Critical Concentration ($C_c$)
As a polymer grows, it depletes the pool of free monomers. Eventually, the monomer concentration drops so low that the rate of addition exactly equals the constant rate of loss. This state of thermodynamic equilibrium (zero net growth) occurs at a specific free monomer concentration known as the Critical Concentration ($C_c$).
- If $[C] > C_c$: The polymer undergoes Net Growth (Elongation).
- If $[C] < C_c$: The polymer undergoes Net Shrinkage (Depolymerization).
Treadmilling: Motion without Growth
Actin filaments possess absolute structural polarity (a fast-growing Plus end and a slow-growing Minus end). Crucially, actin monomers bind ATP. When a monomer is incorporated into the filament, its ATPase activity is triggered. It hydrolyzes ATP to ADP over a delayed time period.
ATP-actin (the "T-form found mostly at the new Plus end) packs tightly and has a very low $C_c$ ($\sim 0.12 \mu M$). Conversely, ADP-actin (the "D-form found mostly at the old Minus end) destabilizes the lattice and has a much higher $C_c$ ($\sim 0.6 \mu M$).
If the cellular free actin concentration is actively maintained between these two values (e.g., exactly at $0.3 \mu M$), a fascinating non-equilibrium steady state is reached. At $0.3 \mu M$, the concentration is high enough to drive addition at the Plus end, but too low to prevent continuous loss at the Minus end.
The polymer maintains a perfectly constant overall length, while individual subunits travel through the filament like a moving escalator—a process named Treadmilling. This unidirectional flux is the primary physical engine driving the leading edge (Lamellipodium) of migrating cells violently forward.
6.4 Microtubules & Dynamic Instability
The explosive collapse of the GTP Cap.
Microtubules are massive, highly rigid hollow cylinders emanating from the Centrosome (MTOC) near the nucleus. Unlike the relatively smooth treadmilling of actin, microtubules exhibit a violently stochastic behavior termed Dynamic Instability. A single microtubule will grow steadily, then suddenly and unpredictably undergo a catastrophic collapse, shrinking rapidly before suddenly transitioning back to growth (Rescue).
The GTP Cap Mechanism
The Stability Physics
The fundamental physics relies on the intrinsic conformational strain of the Tubulin dimer. When bound to GTP, the dimer is perfectly straight, allowing for highly stable lateral bonds between adjacent protofilaments, forming a rigid cylinder.
However, the hydrolysis of GTP to GDP induces a severe conformational bend ($\sim 12^\circ$ curve) in the tubulin dimer. Inside the MT lattice, this GDP-tubulin is forcibly held straight by the surrounding GTP-tubulin at the tip (the GTP Cap), storing massive amounts of elastic mechanical energy like a compressed spring.
The Collapse
The GTPase activity of tubulin is stochastic. If the rate of subunit addition slows down, hydrolysis will eventually catch up to the Plus tip, completely eliminating the protective GTP Cap.
Without the cap to physically restrain them, the GDP-tubulin dimers instantly revert to their preferred curved conformation. The lateral bonds catastrophically fail, and the 13 protofilaments violently peel outward like a banana skin, resulting in extremely rapid depolymerization (shrinking) until the MT is rescued by a new flurry of GTP-tubulin binding.
Clinical Pharmacology: Antimitotic Chemotherapy
During Mitosis (cell division), the cell completely disassembles its interphase microtubule network to construct the massive"
Bind tightly to the inside surface of the $\beta$-tubulin subunit. They hyper-stabilize the microtubule polymer, completely preventing Catastrophe. The spindle freezes, arresting the cancer cell in Metaphase, eventually triggering apoptosis.
Bind to free tubulin dimers in the cytosol, preventing their addition to the Plus end. This guarantees the loss of the GTP cap, forcing the microtubules into continuous, unstoppable depolymerization, obliterating the spindle.
6.6 Molecular Motors
The ATPase Walking Machines.
Microtubules and Actin filaments provide the rigid structural highway network, but they cannot transport cargo alone. The actual movement of massive organelles (like Mitochondria and Vesicles) is executed by sophisticated, ATP-consuming Motor Proteins that convert chemical bond energy into highly directed, step-by-step mechanical force.
Kinesins
+ End DirectedAnterograde Motors. They walk away from the Centrosome (MTOC), transporting cargo outward towards the cell periphery or the axon terminal.
- Kinesin-1 (Conventional) A highly processive dimer. Its two motor "heads operate in a coordinated, strictly alternating hand-over-hand mechanism. The ATP hydrolysis in the rear head triggers a massive conformational swing of the "neck linker," mechanically hurling the other head forward 8nm to bind the next tubulin subunit.
- Kinesin-5 (BimC) A unique bipolar tetramer. It possesses motor heads on both ends. It simultaneously grabs two distinct, antiparallel microtubules and walks towards their Plus ends, actively sliding them apart. This generates the massive pushing force required to separate the Spindle Poles during Mitosis.
Dyneins
- End DirectedRetrograde Motors. They walk back towards the nucleus/MTOC. Structurally, they are completely unrelated to Kinesin or Myosin, being built around a massive, multi-subunit AAA+ ATPase ring complex.
- Cytoplasmic Dynein The massive workhorse for inward traffic. Transports endosomes, lysosomes, and hijacked viruses (e.g., HIV, Rabies) towards the nucleus. Crucially, it cannot bind cargo directly; it strictly requires the giant multi-protein Dynactin Complex (containing p150Glued) to act as an essential tether and processivity factor.
- Axonemal Dynein Highly specialized variants permanently attached to the A-tubules within the 9+2 Axoneme of Cilia and Flagella. When they attempt to walk along the adjacent B-tubule, the structural cross-linking (Nexin) prevents sliding, forcing the entire axoneme structure to undergo rhythmic, wave-like Bending.
6.9 Cell Junctions
Weaving cells into functional tissues.
In multicellular organisms, cells do not exist in isolation. They must structurally adhere to one another and to the extracellular matrix to form tissues capable of withstanding massive physiological stresses (like the pumping of a heart or the stretching of skin). This mechanical integration is achieved via specialized transmembrane protein complexes called Cell Junctions.

Figure 6.2: The primary junctional complexes of simple columnar epithelia.
Tight Junctions(Zonula Occludens)
Located at the very apical edge. Formed by branching strands of Claudin and Occludin proteins. They physically fuse adjacent plasma membranes, acting as an absolute diffusion barrier. They prevent the leakage of extracellular fluids and totally restrict the lateral diffusion of membrane proteins, thus permanently establishing apical/basal cell polarity (essential in the gut lumen).
Adherens Junctions(Zonula Adherens)
Form a continuous adhesion belt just below tight junctions. Transmembrane Classical E-Cadherins bind homophilically to cadherins on the neighbor cell (requiring intense $Ca^{2+}$ concentration). Internally, they are anchored securely to the Actin Cytoskeleton via Catenins. This contractile belt allows epithelial sheets to coordinately fold into tubes during embryogenesis.
Desmosomes(Macula Adherens)
"Spot welds distributed across the lateral membrane, providing immense mechanical resilience. Transmembrane Non-classical Cadherins (Desmoglein/Desmocollin) link cells. Critically, via thick intracellular plaques (Desmoplakin), they anchor to the incredibly tough Intermediate Filament network (Keratin), distributing shear stress across the entire tissue sheet.
Gap Junctions(Nexus)
Not for mechanical strength, but for rapid intercellular communication. Six Connexin proteins form a pore called a connexon. When aligned with a neighbor, they form a continuous 1.5nm hydrophilic channel linking the two cytosols. Allows direct, instant passage of ions ($Ca^{2+}$) and small metabolites ($< 1000$ Da), ensuring synchronous contraction in cardiac muscle.
Module 7:
The Cell Cycle & Apoptosis
7.1 The Control System
The Master Clock: Cyclins and Cdks.
The eukaryotic cell cycle is an irreversible, highly ordered sequence of biochemical events that guarantees the precise duplication of the massive genome and the subsequent physical division of the cell. It is governed by a central biochemical "clock or control system. This master control system is fundamentally constructed from two specific classes of proteins interacting dynamically: Cyclin-Dependent Kinases (Cdks) and their regulatory Cyclin subunits.

Figure 7.0: The Eukaryotic Cell Cycle. A circular series of phases (G1, S, G2, M) controlled by temporal oscillations of Cyclin-Cdk activity.
The Core Engine Components
The catalytic engines. These are master enzymes that continuously phosphorylate downstream target proteins to trigger specific cell cycle events (like nuclear envelope breakdown or DNA replication initiation). Crucially, the intracellular concentration of Cdk proteins remains absolutely constant throughout the entire cell cycle. However, by themselves, they are completely inactive.
The regulatory subunits. They possess no enzymatic activity of their own. They must physically bind to a Cdk to activate it. Furthermore, the specific type of Cyclin completely dictates which target proteins the Cdk will phosphorylate. Unlike Cdks, the concentration of Cyclins violently fluctuates (cycles)—they are cyclically synthesized and then rapidly degraded via targeted proteolysis (ubiquitination) by the APC/C or SCF ligase complexes.
Cyclin Expression Dynamics Over the Cell Cycle
The Major Cyclin-Cdk Complexes
| Complex Name | Cyclin Partner | Cdk Partner | Primary Cellular Function |
|---|---|---|---|
| G1-Cdk | Cyclin D | Cdk4 / Cdk6 | Responds directly to extracellular mitogens. Drives passage through the Restriction Point by heavily phosphorylating the Rb protein, releasing the E2F transcription factor. |
| G1/S-Cdk | Cyclin E | Cdk2 | Irreversibly commits the cell to DNA replication. Prepares the initiation complexes at origins of replication and aggressively induces centrosome duplication. |
| S-Cdk | Cyclin A | Cdk2 | Fires the origins of replication. Crucially phosphorylates Cdc6 and Cdt1 to immediately target them for SCF-mediated destruction, absolutely preventing lethal DNA re-replication. |
| M-Cdk | Cyclin B | Cdk1 | Drives entry into Mitosis. Phosphorylates condensins (for chromosome condensation), lamins (to shatter the nuclear envelope), and MAPs (to organize the bipolar mitotic spindle). |
Fine-Tuning: Switches and Inhibitors
Binding a Cyclin is absolutely necessary, but not always sufficient for full Cdk activation. The cell cycle requires instantaneous, explosive state changes (binary switches), achieved through multi-layered regulation rather than simple gradual accumulation.
- 1. CAK (Cdk-Activating Kinase) Even with Cyclin fully bound, the Cdk active site is partially blocked by a protein loop (the T-loop). CAK must phosphorylate a highly conserved threonine residue on this loop to fully open the active site, achieving maximum catalytic activity.
- 2. Inhibitory Phosphorylation (Wee1 & Cdc25) To stockpile massive amounts of inactive M-Cdk before mitosis, the kinase Wee1 adds inhibitory phosphates to the roof of the active site, completely silencing the complex. To trigger mitosis explosively, the phosphatase Cdc25 removes these inhibitory phosphates. This creates a massive positive feedback loop: active M-Cdk activates more Cdc25 and inhibits Wee1, resulting in an instant, vertical spike in M-Cdk activity.
- 3. CKIs (Cdk Inhibitor Proteins) Proteins like p27, p21, and p16 can physically wrap around the entire Cyclin-Cdk complex, clamping it shut and completely obliterating its kinase activity. This is the primary mechanism used to pause the cell cycle in G1 phase or during severe DNA damage.
7.2 DNA Damage Response (DDR)
The Guardian of the Genome: p53.
If a cell replicates severely damaged DNA or incorrectly separates broken chromosomes, the resulting massive mutations can lead directly to malignant transformation (cancer). Therefore, the cell cycle features rigorous Checkpoints—biochemical surveillance systems that forcefully halt cycle progression until the damage is repaired.
The ATM-p53 Pathway
When DNA suffers catastrophic double-strand breaks (e.g., from ionizing radiation or X-rays), the cell must immediately halt the G1/S transition to prevent replicating the broken DNA template.
-
1. Sensor Detection
The MRN Complex physically detects the broken DNA ends. It immediately recruits and activates the massive apex kinase ATM (Ataxia Telangiectasia Mutated).
-
2. Signal Transduction
Active ATM rapidly phosphorylates a downstream checkpoint kinase called Chk2, massively amplifying the distress signal.
-
3. Stabilization of p53
In a normal, healthy cell, the transcription factor p53 is constantly bound by Mdm2 (an E3 ubiquitin ligase) which continuously destroys p53, keeping its levels extremely low. Chk2 phosphorylates p53. This specific phosphorylation physically blocks Mdm2 from binding. Free from destruction, p53 levels instantly skyrocket.
-
4. Transcriptional Arrest
Stabilized p53 forms a tetramer, enters the nucleus, and powerfully drives the transcription of the CDKN1A gene. This gene produces p21, a potent CKI (Cdk Inhibitor Protein). p21 physically clamps onto G1/S-Cdk and S-Cdk complexes, obliterating their activity. The cell cycle is hard-stopped in G1 until DNA repair enzymes can fix the break.
If p53 is genetically mutated (loss of function), the cell absolutely loses its ability to halt the cycle in response to massive DNA damage. The cell blindly replicates broken chromosomes, rapidly accumulating devastating mutations. p53 is mutated in over 50% of all human cancers, highlighting its indisputable role as the primary Guardian of the Genome.
7.3 The Restriction Point & 7.4 S-Phase
Committing to Division and preventing lethal re-replication.
The Restriction Point (Start) & Rb-E2F
In late G1, mammalian cells encounter the definitive Restriction Point. If a cell successfully passes this point, it is irreversibly committed to completing the entire cell cycle, even if external growth factors (Mitogens) are completely removed from the environment.
- Inhibition: The Retinoblastoma protein (Rb) tightly binds and completely inhibits the E2F transcription factor.
- Signal: Extracellular Mitogens activate the Ras/MAPK pathway, leading to the massive synthesis of Cyclin D, activating G1-Cdk.
- Release: G1-Cdk phosphorylates Rb. Hyperphosphorylated Rb undergoes a massive conformational change and physically releases E2F.
- Transcription: Free E2F enters the nucleus and transcribes S-phase genes (Cyclin E, Cyclin A, DNA polymerase). Crucially, E2F transcribes its own gene, creating an unstoppable positive feedback loop that guarantees irreversible commitment to S-phase.
S-Phase: Licensing and Firing
The mammalian genome contains tens of thousands of Origins of Replication. How does the cell guarantee that every origin fires exactly once per cycle, absolutely preventing lethal re-replication (polyploidy)? The solution is splitting the process into two mutually exclusive biochemical phases separated by Cdk activity.
When Cdk activity is absolutely zero (in G1), the Origin Recognition Complex (ORC) binds the DNA origin. It recruits Cdc6 and Cdt1, which act as loaders to physically clamp the inactive Mcm DNA Helicase double-hexamer around the DNA. This forms the pre-Replicative Complex (pre-RC). The origin is now "Licensed but cannot fire.
The surge in S-Cdk and DDK activity phosphorylates the pre-RC. This physically opens and activates the Mcm helicase, unwinding the DNA to start replication (Firing).
Before the cell can pull sister chromatids apart, it must mathematically guarantee that every single kinetochore is perfectly attached to microtubules from opposite spindle poles (bipolar tension). A single unattached chromosome will result in devastating aneuploidy (e.g., Trisomy 21).
"
The Spindle Assembly Checkpoint (SAC)
The Mechanism of Anaphase
7.7 Apoptosis & Cancer Biology
The ultimate failsafe and its corruption.
If DNA damage is beyond repair, or if a cell receives conflicting proliferative signals, the ultimate failsafe mechanism is engaged: Apoptosis (Programmed Cell Death). This is a highly orchestrated, energy-dependent suicide sequence that dismantles the cell from within without spilling its toxic contents or triggering an inflammatory immune response (unlike necrotic lysis).
Apoptotic Pathways
Driven from within by severe DNA damage or extreme oxidative stress. Relies entirely on catastrophic mitochondrial membrane permeability.
- Pro-apoptotic Bcl2 family proteins (Bax & Bak) oligomerize in the mitochondrial outer membrane, forming massive pores.
- Cytochrome C (normally a crucial electron carrier of the ETC) spills out of the intermembrane space into the cytosol.
- Cytochrome C binds to Apaf-1 adapter proteins, causing them to assemble into a massive 7-spoked wheel of death called the Apoptosome.
- The Apoptosome recruits and activates the initiator Caspase-9, igniting the proteolytic cascade.
Triggered from the outside, usually by a Cytotoxic T-Lymphocyte (Killer T-Cell) executing a dangerous, mutated, or virally infected cell.
- Killer T-Cell displays the Fas Ligand (FasL) on its surface.
- FasL binds to the Fas Death Receptor on the target cell, causing the receptors to cluster and trimerize.
- The cytosolic death domains of Fas recruit adaptor proteins (FADD), forming the massive DISC (Death-Inducing Signaling Complex).
- DISC recruits and activates the initiator Caspase-8, launching the unstoppable cascade.
Convergence: The Executioner Caspases
Both the intrinsic (Caspase-9) and extrinsic (Caspase-8) pathways converge to activate the exact same deadly effector proteases: Executioner Caspases (Caspase-3, -6, -7).
- They ruthlessly cleave Nuclear Lamins, causing total, irreversible collapse of the nuclear envelope.
- They cleave Cytoskeletal Actin and Gelsolin, causing the cell to detach, round up, and undergo severe plasma membrane blebbing.
- They cleave the inhibitor (iCAD) of a latent endonuclease (CAD). CAD is freed, enters the nucleus, and systematically chops the entire genome into precise 200bp fragments (DNA laddering).
Cancer Biology: Corrupting the System
Cancer is fundamentally a disease of a broken cell cycle and failed apoptosis. A single cell requires multiple distinct genetic mutations to become fully malignant, typically affecting two classes of regulatory genes:
Mutated, hyperactive versions of normal genes (proto-oncogenes) that drive cell division. These are Dominant Gain-of-Function mutations; a mutation in only ONE of the two alleles is sufficient to drive cancer.
Examples: Ras (a GTPase permanently stuck in the "ON" state, relentlessly driving the MAPK pathway), Myc (a transcription factor massively overexpressed, forcing entry into S-phase).
Normal genes that pause the cell cycle, repair DNA, or trigger apoptosis. These require Recessive Loss-of-Function mutations; BOTH alleles must be destroyed (Knudson's Two-Hit Hypothesis) for the brake to fail.
Examples: p53 (loss prevents apoptosis in response to massive DNA damage), Rb (loss permanently frees E2F, bypassing the Restriction Point), BRCA1/2 (loss prevents double-strand break repair).
Module 8:
Reproduction & Development
8.1 Meiosis: The Reduction Division
Generating haploid diversity.
Meiosis is a highly specialized cell cycle strictly essential for sexual reproduction. It consists of a single round of DNA replication followed by two successive rounds of chromosome segregation (Meiosis I and Meiosis II). The ultimate objective is to reduce the diploid somatic genome ($2n$) to a haploid state ($1n$) while maximizing genetic diversity through homologous recombination.
Prophase I: The Machinery of Diversity
Prophase I is extraordinarily lengthy and complex, subdivided into five distinct morphological stages to accommodate the physical pairing and recombination of homologous chromosomes.
-
1. Leptotene
Chromosomes begin to condense. The telomeres physically attach to the inner nuclear envelope, grouping together in a highly specific "Bouquet arrangement to facilitate the initial search for their homologous partner in the vast nucleoplasm.
-
2. Zygotene
Homologous chromosomes align perfectly. A massive protein zipper called the Synaptonemal Complex (SC) begins to assemble between them, physically binding them together along their entire length (Synapsis).
-
3. Pachytene (Crossing Over)
Synapsis is complete (forming bivalents/tetrads). A highly conserved endonuclease, Spo11, intentionally induces dangerous double-strand breaks in the DNA. The repair of these breaks utilizing the homologous non-sister chromatid results in reciprocal Crossing Over (Homologous Recombination), physically swapping large chunks of maternal and paternal DNA.
-
4. Diplotene
The Synaptonemal Complex dissolves. The homologous chromosomes attempt to separate but remain physically locked together at the exact sites where crossing over occurred, visibly manifested as X-shaped structures called Chiasmata. (Oocytes arrest in this stage for decades).
-
5. Diakinesis
Maximum chromosome condensation is achieved. The nuclear envelope shatters, and the bivalents prepare to attach to the meiotic spindle for Metaphase I.
Pachytene Crossing Over
8.2 Gametogenesis: Sperm vs Egg
While both processes utilize meiosis, the logistics and resource allocation of Spermatogenesis and Oogenesis are diametrically opposed.
| Feature | Spermatogenesis | Oogenesis |
|---|---|---|
| Product Yield | 4 functional, highly motile spermatozoa. | 1 massive functional Ovum + 3 degenerate Polar Bodies. |
| Timing | Continuous from puberty until death. | Starts in fetal life, arrests for decades, resumes cyclically at puberty. |
| Arrest Points | None. Continuous process. | Arrests in Prophase I (Dictyotene) and Metaphase II. |
| Cytoplasm | Stripped away (Residual Body) to maximize swimming speed. | Conserved completely in the ovum to support early embryonic cleavage. |
The Shugoshin Mechanism
In Anaphase I, Separase cleaves the cohesin along the chromosome arms to resolve the chiasmata, allowing the homologous chromosomes to be pulled apart to opposite poles. However, the sister chromatids must absolutely remain bound together at their centromeres until Anaphase II. How is the centromeric cohesin protected from the rampant Separase enzyme during Anaphase I?
A highly specialized protein aptly named Shugoshin (Japanese for "Guardian Spirit") localizes exclusively to the centromeric region during Meiosis I. Shugoshin physically recruits the PP2A Phosphatase.
While kinases are actively phosphorylating cohesin everywhere else (a prerequisite for Separase cleavage), PP2A constantly removes these phosphates specifically at the centromere. Unphosphorylated cohesin is completely immune to Separase cleavage. Therefore, the sister chromatids remain locked together. In Meiosis II, Shugoshin is intentionally degraded, leaving the centromeric cohesin vulnerable, allowing the final separation of sister chromatids to create haploid gametes.
8.3 Fertilization
The Molecular Handshake & Polyspermy Block.
Fertilization is not a simple random collision; it is a highly orchestrated cascade of biochemical recognition, irreversible membrane fusion, and instantaneous defensive barriers to ensure a strictly diploid zygote is formed.
-
1. Capacitation (The Awakening)
Freshly ejaculated mammalian sperm are completely incapable of fertilizing an egg. As they travel through the female reproductive tract, they undergo Capacitation: massive efflux of cholesterol alters membrane fluidity, and intense calcium influx triggers hyperactivated, whiplash-like motility of the flagellum required to penetrate the cumulus cells.
-
2. The Acrosome Reaction
The sperm forcefully contacts the Zona Pellucida (ZP), a thick glycoprotein matrix encasing the oocyte. Binding to specific ZP receptors triggers massive exocytosis of the sperm's acrosomal vesicle. A payload of powerful hydrolytic enzymes (like hyaluronidase and acrosin) is unleashed, chemically drilling a microscopic tunnel through the ZP shell.
-
3. The Molecular Handshake (Fusion)
Once through the ZP, the plasma membranes of the sperm and egg meet. This crucial fusion is strictly mediated by an indispensable receptor-ligand pair: the sperm presents a transmembrane protein called Izumo1 (named after the Japanese marriage shrine), which irreversibly binds to an essential receptor on the egg surface called Juno (the Roman goddess of marriage). Only upon this specific binding do the lipid bilayers merge, allowing the sperm nucleus to enter the oocyte cytoplasm.
The Cortical Reaction (Polyspermy Block)
If a second sperm fertilizes the egg (Polyspermy), the resulting triploid embryo will suffer catastrophic mitotic spindle failure and die immediately. The egg must deploy an absolute, impenetrable block within seconds of the first fusion.
The fusion of Izumo1 and Juno triggers an explosive release of $Ca^{2+}$ from the egg's endoplasmic reticulum. This creates a massive calcium wave that visibly sweeps across the entire oocyte. This calcium surge triggers the immediate exocytosis of thousands of Cortical Granules located just beneath the plasma membrane.
These granules dump enzymes into the space beneath the Zona Pellucida. These enzymes aggressively cleave the ZP proteins (ZP2), permanently hardening the shell into an impenetrable concrete barrier. Simultaneously, all remaining Juno receptors are completely shed from the egg surface, ensuring no other sperm can ever bind.
8.5 Stem Cells & iPSCs
Epigenetic Reprogramming.
Embryonic Stem Cells (ESCs)
Derived from the Inner Cell Mass (ICM) of a 5-day-old pre-implantation blastocyst. These cells are truly Pluripotent: they possess the unrestricted ability to differentiate into every single cell type of the adult body (all three germ layers: ectoderm, mesoderm, endoderm), but they cannot form the extra-embryonic placenta (which is derived from the outer trophoblast).
Driven by three master transcription factors: Oct4, Sox2, and Nanog. They form an auto-regulatory positive feedback loop to maintain stemness and aggressively suppress all differentiation genes.
Induced Pluripotent Stem Cells (iPSCs)
A revolutionary breakthrough in regenerative medicine (Shinya Yamanaka, Nobel Prize 2012). It was proven that terminally differentiated, specialized adult somatic cells (like skin fibroblasts) could be forced to reverse time and return to an embryonic-like pluripotent state.
Retroviral introduction of just four specific transcription factors: Oct4, Sox2, Klf4, and c-Myc. These factors aggressively rewrite the epigenetic landscape, wiping out the somatic methylation marks and re-establishing the open chromatin structure of an ESC, bypassing the severe ethical dilemmas of harvesting human embryos.
Module 9:
Preservation Technology
9.3 Cryobiology Thermodynamics
Mazur's Two-Factor Hypothesis.
The objective of cryopreservation (storage in liquid nitrogen at -196°C) is to halt all biological time by arresting molecular kinetics. However, freezing water is inherently lethal to cells. The challenge of cryobiology is defined by Peter Mazur's definitive Two-Factor Hypothesis: cell survival dictates finding the absolute "Goldilocks cooling rate, balancing two opposing vectors of cellular destruction.
Survival vs. Cooling Rate
Factor 1: Cooling Too Slowly (The Solution Effect)
If cooled too slowly, ice crystals form exclusively in the extracellular space first. As pure water freezes out, the remaining unfrozen extracellular fluid becomes a hyper-concentrated, toxic salt brine. Massive osmotic pressure forces water completely out of the cell. The cell undergoes severe, fatal dehydration, collapsing structurally, while the soaring salt concentration irreversibly denatures internal proteins.
Factor 2: Cooling Too Fast (Intracellular Ice)
If cooled too rapidly (e.g., dropping a vial straight into LN2 without cryoprotectants), there is no time for osmotic dehydration to occur. The water is trapped inside the cell and freezes violently in place. The expanding, razor-sharp Intracellular Ice Formation (IIF) instantly punctures and shreds the plasma membrane and internal organelles. This is 100% lethal.
The Optimal Rate (The Compromise)
Maximum survival requires a highly controlled cooling rate (typically ~1°C per minute). This precise speed provides just enough time for water to safely exit the cell (preventing internal ice spikes) but is fast enough to minimize the duration the cell is exposed to the toxic, hypertonic "Solution Effect brine.
Cryoprotectants (CPAs)
Even with the perfect cooling rate, survival is impossible without chemical intervention. Cells must be incubated in Cryoprotective Agents (CPAs) before freezing.
Small molecules that pass directly through the plasma membrane. They physically replace the water inside the cell. They possess extreme hydrogen-bonding capacity, which dramatically lowers the freezing point and severely disrupts the rigid geometric lattice of ice crystals, forcing the water to freeze into a harmless, amorphous slush rather than sharp shards.
DMSO is highly cytotoxic at room temperature. Upon thawing (which must be rapid), the cells must be immediately washed or diluted into massive volumes of media to remove the DMSO before it triggers apoptosis.
9.4 Vitrification (Glass Formation)
Vitrification completely abandons the slow-cooling paradigm. The objective is to drop the temperature so instantaneously that water molecules literally do not have the kinetic time to organize into an ice crystal lattice. Instead, the liquid phase undergoes a profound physical transition directly into a solid, amorphous Glass state.
- Ultra-Fast Cooling Rates: Searing velocities up to 20,000°C/min. Achieved by plunging samples directly into Slush Nitrogen (-210°C).
- Massive CPA Concentration: Requires toxic levels of CPAs (up to 40-60% v/v) to artificially spike the viscosity of the solution, mechanically freezing molecules in place before they can crystallize.
- Micro-Volumes: Utilizing microscopic carriers (like Cryotops or EM grids) holding less than 1µL of media to eliminate the Leidenfrost effect (nitrogen boiling gas barrier) and guarantee instantaneous heat transfer.
Highly specialized for immense, water-rich structures exquisitely sensitive to ice damage, specifically Human Oocytes and Pre-implantation Embryos.