IMAT Biomolecules I: Carbs & Lipids
🔬 0. Introduction to Macromolecules
Biological macromolecules are massive, highly complex organic molecules that construct the physical architecture and the functional, catalytic machinery of all living organisms. The vast majority of these macromolecules are polymers—long, repeating chains assembled from smaller, simpler molecular building blocks known as monomers. The process of linking these monomers together is one of the most fundamental reactions in all of biochemistry.
The Universal Chemistry of Polymerization
Regardless of whether the cell is synthesizing a carbohydrate, a protein, or a nucleic acid, the chemical mechanisms underlying the assembly and disassembly of these polymers revolve entirely around the molecule of Water ($H_2O$).
This is an anabolic process. Two separate monomers are covalently joined together to form a larger polymer. In the process, a hydroxyl group ($-OH$) is removed from one monomer, and a hydrogen atom ($-H$) is removed from the other. These immediately combine to form and release a molecule of water.
Because building complex, highly ordered bonds from free-floating monomers decreases the entropy of the system, this process is strictly endergonic and always requires an input of energy (usually supplied by the hydrolysis of ATP).
This is a catabolic process. It is the exact chemical reverse of condensation. To break the covalent bond holding a polymer together, a molecule of water is forcefully inserted (lysed) across the bond, restoring the $-OH$ and $-H$ to the respective individual monomers, thereby separating them.
This process increases entropy and is therefore highly exergonic, releasing the energy stored in the chemical bond. It is the fundamental biochemical mechanism behind all food digestion in the human gastrointestinal tract.
🍞 1. Carbohydrates (Saccharides): Monosaccharides
Carbohydrates are polyhydroxy aldehydes or ketones. Historically named because their chemical formulas make them appear as "hydrates of carbon", they are composed exclusively of Carbon, Hydrogen, and Oxygen atoms, almost universally obeying the strict stoichiometric ratio of $1:2:1$. This is represented by the empirical formula $(CH_2O)_n$. They are the most abundant class of biomolecules on Earth, acting as the primary fuel source for cellular respiration and as robust structural components.

Monosaccharides (Simple Sugars)
Monosaccharides are the simplest, indivisible monomeric units of carbohydrates. Due to the presence of multiple highly polar, electronegative hydroxyl ($-OH$) functional groups along their carbon chain, they form extensive hydrogen bonds with water. This makes them extremely soluble in aqueous solutions, forming sweet-tasting, crystalline solids at room temperature. They are classified biochemically by two primary metrics: the total number of carbon atoms they possess, and whether their defining carbonyl group ($C=O$) is an aldehyde (located at the end of the chain) or a ketone (located internally).
| Class Name | Carbons | Chemical Formula | Key Biological Examples & Functions |
|---|---|---|---|
| Triose | 3 | $C_3H_6O_3$ | Glyceraldehyde (G3P): An absolutely crucial 3-carbon intermediate metabolite found universally in both cellular respiration (specifically within glycolysis) and photosynthesis (as the direct product of the Calvin Cycle). |
| Pentose | 5 | $C_5H_{10}O_5$ |
Ribose: The structural 5-carbon backbone of RNA, ATP, and essential electron carriers like $NAD^+$. It possesses a critical, reactive hydroxyl group ($-OH$) at the Carbon-2 position. Deoxyribose ($C_5H_{10}O_4$): The structural backbone of DNA. As the name heavily implies ("De-oxy"), it lacks the oxygen atom at the Carbon-2 position, containing only a Hydrogen atom there instead. This lack of a highly reactive oxygen makes the DNA molecule vastly more chemically stable and resistant to hydrolysis than RNA. |
| Hexose | 6 | $C_6H_{12}O_6$ |
Glucose (Aldose): The primary blood sugar in humans and the universal fuel for cellular aerobic respiration. Fructose (Ketose): Fruit sugar. The sweetest naturally occurring sugar, processed primarily in the liver. Galactose (Aldose): A structural component of mammalian milk sugar (lactose). It cannot be directly utilized for energy by the brain; it must first be enzymatically converted to glucose in the liver. |
Deep Dive: Isomerism & Molecular Chirality
Because biological enzymes are staggeringly specific 3D protein structures, the exact spatial orientation of atoms in a substrate is just as important as its raw chemical formula. Monosaccharides exhibit multiple, overlapping layers of complex isomerism.
Epimers are a highly specific type of diastereomer that differ in spatial configuration at exactly one specific chiral carbon. For instance, D-Glucose and D-Galactose are C-4 epimers. They are structurally identical everywhere except that the $-OH$ group on Carbon-4 points in the opposite spatial direction. This tiny geometric flip makes Galactose completely unrecognizable to the enzyme Hexokinase.
The new $-OH$ group on this anomeric carbon can lock into one of two distinct positions relative to the bulky $CH_2OH$ group at C-6:
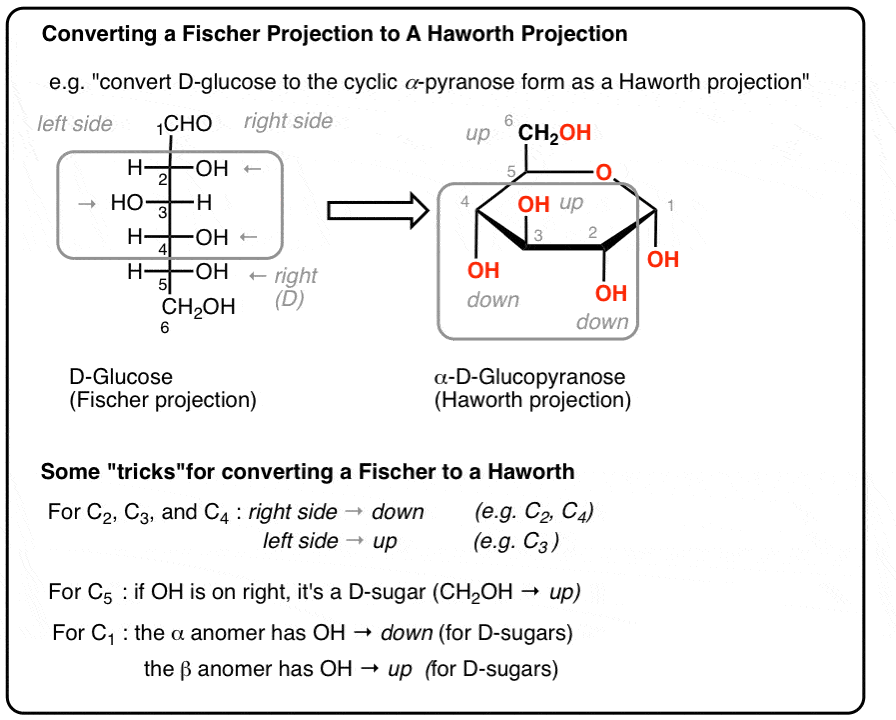
Structural Visualization: The Anomers of Glucose
🍩 2. Disaccharides: Double Sugars
Disaccharides are constructed by the enzymatic, endergonic condensation (dehydration synthesis) of two specific monosaccharides. The resulting strong covalent bond holding the two sugars together is called a Glycosidic Bond. Like all condensation reactions, the formation of this bond results in the loss of one molecule of water.
Mechanism: Glycosidic Bond Formation
The most common linkage in biology occurs between the highly reactive anomeric carbon (Carbon-1) of the first sugar molecule and the relatively stable Carbon-4 of the second sugar molecule. This specific geometry creates a linear 1 $\rightarrow$ 4 Glycosidic Linkage.
Biochemical Mechanism: Condensation Synthesis of Maltose
The "Big Three" Disaccharides (High Yield)
| Disaccharide Name | Monomers & Bond Type | Properties & Biological Significance |
|---|---|---|
| Maltose (Malt Sugar) |
$\alpha$-Glucose + $\alpha$-Glucose $\alpha(1 \rightarrow 4)$ |
Produced in massive quantities during the digestion of starch by the enzyme Amylase (found in saliva and pancreatic juice). It retains a free anomeric carbon, making it a highly reactive Reducing Sugar. |
| Sucrose (Table Sugar) |
$\alpha$-Glucose + $\beta$-Fructose $\alpha(1 \rightarrow 2)\beta$ |
The standard table sugar extracted from sugar cane. It is the primary transport sugar in the phloem sap of plants precisely because the unique bond locks both anomeric carbons, rendering it exceptionally stable and unreactive (Non-Reducing Sugar). |
| Lactose (Milk Sugar) |
$\beta$-Glucose + $\beta$-Galactose $\beta(1 \rightarrow 4)$ |
The primary sugar found in mammalian milk, providing dense energy for nursing infants. Its beta bond makes it difficult to digest, requiring a highly specific enzyme to break. It is a Reducing Sugar. |
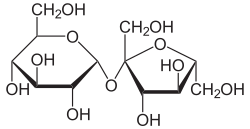
A "reducing sugar" is chemically defined as any carbohydrate capable of acting as a reducing agent. To do this, the sugar must possess a "free" anomeric carbon capable of momentarily opening its ring structure in aqueous solution to expose a highly reactive aldehyde or ketone functional group.
In maltose and lactose, the glycosidic bond only utilizes Carbon-1 of the first sugar. The anomeric carbon on the second sugar remains untouched and "free" to open, react, and reduce other chemicals (like the copper in Benedict's reagent).
The Sucrose Exception: In Sucrose, the condensation reaction occurs directly between Carbon-1 of glucose AND Carbon-2 of fructose. Because both of these are the anomeric carbons of their respective rings, both rings are permanently locked shut by the bond. Neither can open to form an aldehyde/ketone. Therefore, sucrose is chemically inert and is termed a Non-Reducing Sugar.
Lactose Intolerance
Mammals synthesize the enzyme Lactase as infants specifically to cleave the stubborn $\beta(1 \rightarrow 4)$ glycosidic bond of lactose. In most humans worldwide (except populations with a specific evolutionary mutation), lactase gene expression plummets after weaning. If a lactase-deficient adult consumes dairy, the intact, bulky lactose travels undigested into the large intestine.
Pathology: The heavy lactose molecules wildly increase the osmolarity of the colon, drawing massive amounts of water out of the blood and into the gut via osmosis, causing severe, explosive diarrhea. Simultaneously, opportunistic anaerobic gut bacteria rapidly ferment the lactose, producing excessive $CO_2$, methane, and hydrogen gas, leading to painful bloating and flatulence.
Galactosemia
A severe, potentially fatal autosomal recessive genetic metabolic disorder. Infants born with this disease lack the specific enzymes (most commonly GALT) required to convert the galactose (derived from the breakdown of milk lactose) into usable glucose.
Pathology: Toxic levels of galactose and its heavily reduced, highly osmotic metabolite (galactitol) rapidly accumulate in the blood and tissues. This buildup causes catastrophic liver damage (hepatomegaly and cirrhosis), profound mental retardation, and the rapid, irreversible formation of bilateral cataracts in the eyes of newborns within days of drinking milk.
🌳 3. Polysaccharides: Complex Polymers
Polysaccharides are massive, towering macromolecules composed of hundreds to many thousands of monosaccharides linked by glycosidic bonds. Unlike simple sugars, they are entirely insoluble in water (meaning they are osmotically neutral and do not draw water into the cell, which is absolutely crucial for safe intracellular storage) and they are completely tasteless. Their macroscopic biological function—whether they act as a cellular battery (energy storage) or as physical building materials (structural support)—is determined entirely by the specific isomer of glucose they are built from ($\alpha$ vs $\beta$) and the specific geometry of their glycosidic branching.
A. Energy Storage Polymers
(Built exclusively from $\alpha$-Glucose)
Plants store their excess glucose manufactured during photosynthesis as dense starch granules within specialized plastids called amyloplasts. Starch is a heterogeneous mixture of two distinct polymers:
- Amylose (approx. 20%): An unbranched, perfectly linear chain. Consists exclusively of $\alpha(1 \rightarrow 4)$ bonds. Due to bond angles, it naturally coils into a tight helical spring, making it incredibly compact but slow to digest (enzymes can only attack the two ends).
- Amylopectin (approx. 80%): A highly complex, branched polymer. Contains the $\alpha(1 \rightarrow 4)$ backbone plus $\alpha(1 \rightarrow 6)$ branch points. Faster hydrolysis due to numerous terminal branches.
Stored in massive, dense granules primarily within the hepatocytes of the Liver (to maintain blood sugar levels) and inside Skeletal Muscle cells. Structurally, it is nearly identical to amylopectin, but it is vastly more highly branched, with $\alpha(1 \rightarrow 6)$ branch points occurring rapidly every 8-12 glucose units.
This extreme branching creates tens of thousands of terminal ends on a single molecule. When adrenaline spikes, enzymes (glycogen phosphorylase) attack all these ends simultaneously, flooding the bloodstream with a massive, instantaneous tsunami of glucose to power the "Fight or Flight" response.
B. Structural Support Polymers
(Built exclusively from $\beta$-Glucose)
Cellulose is the primary structural component of the tough, rigid cell walls that enclose plant cells. It is, by mass, the single most abundant organic polymer on Earth. Humans completely lack the enzymes to digest cellulose, making it the primary component of dietary fiber.
Because the $-OH$ group on C1 of $\beta$-glucose points UP, forming a bond requires every alternating monomer to physically flip upside down ($180^\circ$) relative to its neighbor. This prevents coiling into a helix, creating perfectly straight chains.
When dozens of these straight chains lie parallel to each other, the exposed $-OH$ groups form massive networks of Hydrogen Bonds between the parallel chains. These cross-linked cables are called Microfibrils, granting wood its immense tensile strength.
Chitin is a vital structural polysaccharide utilized by arthropods (insects, spiders, crustaceans) to build their tough, flexible exoskeletons, and by fungi to build their cell walls. It utilizes the exact same $\beta(1 \rightarrow 4)$ linkages and $180^\circ$ alternating flips as cellulose. However, its monomer is a modified sugar called N-acetylglucosamine, which possesses a nitrogen-containing appendage that vastly increases hydrogen bonding potential, making the exoskeleton extremely tough, leathery, and waterproof.
Architectural Schematic: Biological Polysaccharides
Diabetes Mellitus & Glycogen Storage Diseases
A systemic metabolic disorder characterized by chronic hyperglycemia. Glucose is highly osmotically active, drawing water out of cells, leading to severe dehydration, polyuria, and polydipsia.
- Type 1 (Autoimmune): Destruction of pancreatic beta cells. Zero endogenous insulin produced.
- Type 2 (Resistance): Cells ignore the insulin signal due to receptor down-regulation. Strongly linked to central obesity.
- HbA1c Diagnostic Test: Glucose spontaneously and permanently binds to hemoglobin in RBCs (glycation). Measuring HbA1c provides a perfectly accurate, un-cheatable 3-month average of blood sugar levels.
Rare genetic disorders where specific enzymes required for the synthesis or breakdown of glycogen are defective, leading to toxic accumulation of abnormal glycogen in tissues.
- Von Gierke's Disease (Type I): Deficiency in Glucose-6-Phosphatase. The liver cannot release free glucose into the blood. Causes severe fasting hypoglycemia and massive hepatomegaly.
- Pompe's Disease (Type II): Lysosomal enzyme deficiency. Glycogen destroys muscle cells, particularly the heart, causing fatal cardiomegaly in infants.
🧪 4. Analytical Biochemistry: Lab Tests for Carbohydrates
Biological sciences heavily rely on qualitative and semi-quantitative chemical reagents to detect the presence of specific macromolecules in unknown solutions. The exact biochemical physical chemistry theory behind these color-change reactions is a frequent target for high-level IMAT questions.
Benedict's Test
(Reducing Sugars)
Chemical Theory: Benedict's reagent contains Copper (II) Sulfate ($Cu^{2+}$), which is a brilliant, clear blue in solution. When boiled with a "reducing sugar" (like Glucose or Maltose), the free aldehyde/ketone group of the sugar acts as a strong reducing agent. It physically donates an electron to the Copper ion, reducing the blue $Cu^{2+}$ into $Cu^+$. The $Cu^+$ precipitates out of the aqueous solution as insoluble Copper (I) Oxide ($Cu_2O$), which is a heavy, opaque Brick-Red solid.
Semi-Quantitative Colorimetric Scale
(Blue)
(Green)
(Yellow)
(Orange)
(Brick Red)
Iodine Test
(Detection of Starch)
Chemical Theory: Potassium Iodide ($I_2/KI$ solution) is normally a clear, pale orange-brown liquid. The unique structural geometry of Amylose (the unbranched component of plant starch) forms a tightly coiled, hydrophobic helix. The triiodide ion complexes ($I_3^-$) are the exact perfect physical size and shape to slip inside the hollow core of this amylose helix. When trapped inside, an intense electron charge-transfer complex is formed between the starch and the iodine, completely altering how the molecule absorbs light.
🥑 5. Lipids & Fatty Acid Foundations
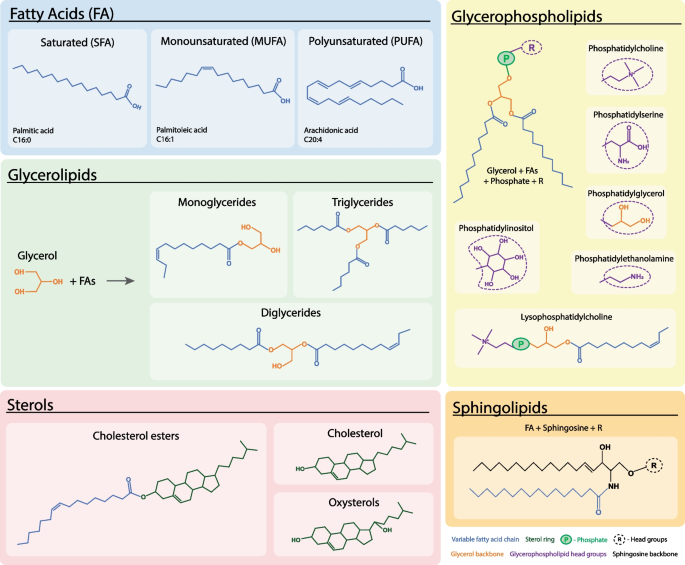
Lipids are an exceptionally diverse group of biological molecules that are grouped together not by a shared structural monomer (like all other macromolecules), but by a shared physical property: they are entirely or almost entirely hydrophobic (insoluble in aqueous solutions like water or blood). However, they dissolve readily in non-polar organic solvents like ethanol, chloroform, or ether. They contain Carbon, Hydrogen, and Oxygen, but with a drastically lower proportion of Oxygen atoms compared to carbohydrates, making them highly reduced.

The Fundamental Building Blocks: Fatty Acids
A fatty acid consists of a highly acidic, polar, hydrophilic carboxyl functional group ($-COOH$) attached firmly to a long, completely non-polar, hydrophobic hydrocarbon chain (the R-group tail). The length of this hydrocarbon tail (typically 12-24 carbons) and the specific presence or absence of double bonds definitively determine the physical properties (like melting point) of the fat.
| Fatty Acid Type | Chemical Structure ($C-C$ bonds) | Physical Properties | Primary Dietary Sources |
|---|---|---|---|
| Saturated | Contains absolutely zero double bonds between carbon atoms. Every single carbon is fully "saturated" with the maximum theoretically possible number of covalently bonded Hydrogen atoms. | Solid (at room temp $20^\circ C$) Straight chains pack tightly together, maximizing strong intermolecular Van der Waals forces. |
Animal fats (lard, tallow, bacon fat), Butter, Cheese, Coconut Oil. |
| Unsaturated (Cis configuration) |
Contains one (monounsaturated) or multiple (polyunsaturated) $C=C$ double bonds within the hydrocarbon backbone. | Liquid / Fluid (at room temp $20^\circ C$) Cis double bonds create rigid $30^\circ$ kinks, severely preventing tight physical packing and drastically lowering the melting point. |
Plant-derived oils (Olive oil, sunflower oil), Fish oils (Salmon), Avocados, Nuts. |
Molecular Geometry: Saturated vs. Unsaturated Architectures
Essential Fatty Acids & The Danger of Trans Fats
Mammals lack the specific desaturase enzymes required to introduce double bonds beyond carbon-9 in a fatty acid chain. Therefore, certain highly polyunsaturated fats must be consumed entirely through diet. These are termed "Essential".
The "Omega" number simply indicates the position of the very first double bond, counting backwards starting from the methyl ($CH_3$) end (the omega end) of the chain, rather than the carboxyl ($COOH$) alpha end.
- Omega-3 ($\alpha$-linolenic acid): Highly anti-inflammatory precursors. Critical for brain/retina development and synthesizing anti-inflammatory eicosanoids. Sources: Fatty fish (Salmon), flaxseeds, walnuts.
- Omega-6 (Linoleic acid): Precursors to pro-inflammatory eicosanoids (prostaglandins, leukotrienes). Essential, but massive overconsumption in modern western diets drives chronic systemic inflammation. Sources: Cheap vegetable seed oils.
Trans fats are overwhelmingly artificial, created by the industrial food process of partial hydrogenation (bombarding cheap, liquid vegetable oils with hydrogen gas at extreme high heat to make them solid and shelf-stable, like margarine and shortening).
This violent process chemically flips the natural cis double bonds into unnatural trans double bonds. A trans double bond removes the kink! It physically straightens the hydrocarbon chain, forcing an unsaturated fat to behave physically exactly like a saturated fat (solid at room temp).
Cardiovascular Pathology: Human lipase enzymes cannot properly digest trans fats. They radically increase LDL (bad cholesterol), plummet HDL (good cholesterol), and trigger massive systemic inflammation and rapid atherosclerosis (arterial plaque buildup). They are effectively toxic.
🥓 6. Triglycerides (Triacylglycerols)
Triglycerides are the primary, most highly concentrated form of long-term energy storage in the biological world, found in the adipose tissue of animals and the seeds/oils of plants. Because they are entirely non-polar and hydrophobic, they are stored in a completely anhydrous (water-free) state inside cells, making them incredibly lightweight and space-efficient compared to water-logged glycogen.
Biochemical Formation (Esterification)
A triglyceride is synthesized via a massive triple-condensation reaction between exactly 1 molecule of Glycerol (a 3-carbon alcohol backbone) and 3 molecules of Fatty Acids. This reaction forms three covalent Ester Bonds. Because it is a 3-part condensation, the synthesis of one single triglyceride molecule forcibly releases exactly 3 molecules of Water ($H_2O$).
Saponification is the alkaline hydrolysis of the ester bonds in a triglyceride. When a triglyceride is boiled with a strong base (like $NaOH$ or $KOH$), the ester bonds are cleaved, releasing free glycerol and the sodium/potassium salts of the fatty acids, which we universally call Soap. These amphipathic soap molecules can surround dirt and oil, forming micelles that wash away in water.

Reaction Mechanism: Esterification & Triglyceride Assembly
Physiological Functions of Triglycerides
Yields 9 kcal/g (38 kJ/g), more than double the energy density of carbohydrates or proteins. Because it perfectly repels water, it takes up vastly less physical volume and weight in an organism compared to heavily hydrated glycogen.
Adipose tissue acts as a superb thermal insulator against severe cold. Marine mammals (whales, seals, walruses) rely heavily on extremely thick layers of subcutaneous fat called blubber to survive in freezing ocean temperatures.
Visceral fat deposits form protective, shock-absorbing cushions around vital internal organs, most notably the delicate kidneys and the heart, protecting them from severe physical trauma and impact.
The complete aerobic oxidation of lipid hydrocarbon chains in the mitochondria yields a massive amount of $H_2O$ as a byproduct. Desert animals (like the Camel's hump) store immense amounts of fat specifically to be burned to generate vital metabolic water during severe droughts.
🌊 7. Phospholipids, Sphingolipids & Membranes
Phospholipids (specifically glycerophospholipids) are the absolute fundamental architectural basis of all biological membranes on Earth. They are created when one of the three fatty acid tails of a standard triglyceride is removed and replaced by a highly negatively charged, bulky Phosphate Group. This singular chemical substitution completely revolutionizes the physics of the molecule, transforming it from a simple, entirely hydrophobic fat droplet into a highly sophisticated, two-faced Amphipathic molecule.
- The "Head" Region: Composed of the glycerol backbone, the phosphate group, and often an additional polar organic molecule attached to the phosphate (like choline, ethanolamine, or serine). Because the phosphate carries a heavy negative charge, the head is extremely polar and fiercely Hydrophilic (Water-loving).
- The "Tail" Region: Composed of the two remaining long hydrocarbon fatty acid chains. These chains are completely non-polar, possess zero electrical charge, cannot form hydrogen bonds, and are therefore fiercely Hydrophobic (Water-fearing). Usually, one tail is straight (saturated) and the other is kinked (unsaturated), which precisely dictates the physical fluidity of the membrane.
Not all membrane lipids use a glycerol backbone. Sphingolipids use a complex amino alcohol called sphingosine as their backbone. The most famous example is Sphingomyelin, which is heavily concentrated in the Myelin Sheath that wraps around and electrically insulates the axons of neurons in the central and peripheral nervous systems, allowing for saltatory conduction of action potentials.
Clinical Note: Multiple Sclerosis (MS) is an autoimmune disease where the immune system destroys this sphingomyelin-rich sheath, crippling nerve transmission. Additionally, a genetic defect in the lysosomal breakdown of sphingolipids leads to catastrophic accumulation diseases, such as Tay-Sachs disease.
When millions of these amphipathic molecules are dumped into an aqueous environment (like the watery interior and exterior of a human cell), the absolute laws of physics and thermodynamics force them to spontaneously self-assemble. To perfectly hide their hydrophobic tails from the water while simultaneously exposing their hydrophilic heads to the water, they instantly form a highly stable, self-sealing double layer known as a Phospholipid Bilayer. This dynamic bilayer is the matrix of the Fluid Mosaic Model of cell membranes.
Architectural Schematic: The Phospholipid Bilayer

Lipoproteins: The Taxis of the Bloodstream
Blood is overwhelmingly composed of water. Lipids (triglycerides and cholesterol) are strictly hydrophobic; if dumped directly into the blood from the liver or intestines, they would instantly coalesce into massive, lethal fat embolisms that would clog arteries and stop the heart. To safely transport lipids through the aqueous circulatory system, the body packages them into highly complex, water-soluble spheres called Lipoproteins.
A lipoprotein is structured like a biochemical submarine: It features an outer monolayer composed of amphipathic phospholipids and special identifying structural proteins called Apolipoproteins (the hydrophilic heads face the blood). The hollow interior core is stuffed entirely full of highly hydrophobic triglyceride cargo and cholesterol esters.
Clinically termed the "Bad Cholesterol". LDL acts as a delivery truck, transporting cholesterol manufactured in the liver out into the peripheral tissues and blood vessels. If a patient has excess LDL, it aggressively invades the endothelial walls of arteries, undergoes rapid oxidation, and gets eaten by macrophages. These macrophages swell and die, forming a fatty streak that hardens into Atherosclerotic Plaque, massively increasing the risk of myocardial infarctions (heart attacks) and strokes.
Clinically termed the "Good Cholesterol". HDL acts as the body's vascular garbage truck. It constantly circulates through the bloodstream, physically scavenging excess, rogue cholesterol from the peripheral tissues and artery walls, actively pulling it out of developing plaques. It safely transports this dangerous cholesterol back to the liver, where it is metabolized into bile acids and permanently excreted from the body via the intestines (Reverse Cholesterol Transport).
💍 8. Steroids & Waxes
Despite having absolutely zero physical or structural resemblance to triglycerides or phospholipids (they contain no fatty acid chains whatsoever), Steroids are formally classified as lipids exclusively because they are fiercely hydrophobic and completely insoluble in water.
Steroids in Human Biology
All steroid hormones in the human body are synthesized directly from a single master precursor molecule: Cholesterol. Because steroid hormones are uniquely lipid-soluble, they do not bind to receptors on the outside of target cells like bulky peptide hormones do. Instead, they slip effortlessly straight through the hydrophobic plasma membrane, entering the cytoplasm or nucleus directly to alter gene transcription at the DNA level.
- Cholesterol (The Membrane Buffer): Synthesized primarily in the liver by the enzyme HMG-CoA Reductase (the enzyme targeted by Statin drugs to lower cholesterol). Besides making hormones, it physically wedges itself between the phospholipids of animal cell membranes. It acts as a bidirectional "fluidity buffer": at high temperatures, it physically restricts phospholipid movement, preventing the membrane from melting. At freezing temperatures, its bulky structure forces the phospholipids apart, preventing them from packing tightly and freezing solid.
- Corticosteroids: Synthesized by the adrenal cortex. Includes Cortisol (the primary long-term stress hormone, which is heavily immunosuppressive and stimulates gluconeogenesis) and Aldosterone (regulates blood pressure by controlling sodium reabsorption in the kidneys).
- Sex Hormones: Synthesized primarily by the gonads. Includes Testosterone, Estrogen, and Progesterone. Responsible for sexual differentiation, secondary sex characteristics, and regulating the menstrual cycle and pregnancy.
Waxes
Waxes are highly complex, extremely stable esters formed by the condensation of an incredibly long-chain fatty acid with an equally long-chain complex alcohol. Because their hydrocarbon chains are so massive, they are the most fiercely hydrophobic biological molecules in existence, solid at room temperature.
Biological Role: They are utilized almost exclusively for heavy-duty waterproofing and protection. Plants secrete dense layers of wax over their leaves (the cuticle) to prevent lethal water loss via transpiration in the hot sun. Bees use beeswax to construct the intricate structural architecture of their honeycombs. Birds and waterfowl coat their feathers in wax secreted from a preen gland to remain buoyant and dry.
🧪 9. Analytical Biochemistry: Lab Tests for Lipids
Testing for lipids is fundamentally different from testing for carbohydrates or proteins. Because lipids lack reactive functional groups (they are essentially just inert hydrocarbon chains), we cannot use a simple colorimetric chemical reaction (like Benedict's or Biuret). Instead, the standard laboratory test for lipids relies entirely on their defining physical property: their strict insolubility in water.
The Emulsion Test
Physical Chemistry Theory: Lipids are completely non-polar, meaning they cannot dissolve in polar water. However, they dissolve very easily in non-polar or moderately polar organic solvents, such as pure absolute Ethanol. By forcing the lipid to dissolve in ethanol first, and then violently shocking the solution by pouring it into a large volume of cold water, the lipid molecules are forcefully ripped out of solution. Because they cannot interact with the water, they instantly precipitate, clumping together to form millions of microscopic, insoluble fat droplets suspended in the water. This suspension of droplets is called an emulsion.
Strict Methodology:- Take a solid or liquid food sample and place it in a completely dry, grease-free test tube.
- Add 5ml of absolute, concentrated Ethanol to the tube.
- Shake the tube violently for 1-2 minutes. (This physically extracts and fully dissolves any lipids present in the sample into the ethanol).
- Decant (pour) the liquid portion containing the dissolved lipid into a second, clean test tube filled with cold, distilled Water.
-
Positive Result: The immediate formation of a thick, cloudy, opaque White Emulsion near the top of the water.
Physics Note: The emulsion appears milky white due to the Tyndall effect—the millions of tiny suspended lipid droplets physically scatter the light hitting the tube.
Visual Result
Cloudy White Emulsion
⚡ 10. Thermodynamics: Energy Yield & Respiratory Quotient
A frequent, high-yield IMAT question asks to compare the physical and chemical properties of carbohydrates versus lipids as energy storage molecules. Why does a mobile animal store the vast bulk of its energy as fat rather than just building a massive brick of glycogen?
| Biochemical Feature | Carbohydrates (Glycogen) |
Lipids (Triglycerides) |
|---|---|---|
| Physiological Use Case | Short-Term, Rapid Energy. (Burned during sprinting or immediate stress. Soluble, easy to mobilize). | Long-Term, Dense Energy. (Burned during resting, endurance, or starvation). |
| Total Energy Density | ~17 kJ/g (4 kcal/g) |
~38 kJ/g (9 kcal/g) |
| Deep Chemistry Reason | Carbohydrates are already partially oxidized. They contain a massive amount of heavy Oxygen atoms ($-OH$ groups) relative to Carbon. Because they are already partially oxidized, completing their combustion yields significantly less net energy. | Lipids are highly, heavily reduced. They consist almost entirely of non-polar C-H bonds with virtually zero oxygen atoms. Oxidizing these pure hydrocarbon chains in the mitochondria yields vast quantities of high-energy electrons for the ETC. |
| Physical Storage Weight | Carbohydrates are intensely hydrophilic. In cells, 1 gram of glycogen chemically binds to and traps about 2-3 grams of heavy Water. Storing massive amounts of energy as glycogen would make an animal far too heavy and bulky to walk or escape predators. | Lipids are fiercely hydrophobic. They are stored completely anhydrously (without a single drop of water) in dense fat droplets inside adipocytes. This extreme weight efficiency is an absolute evolutionary mandate for mobile animals and flying birds. |
The RQ is a dimensionless number used in basal metabolic rate calculations to determine exactly which macronutrient a patient or organism is currently burning for energy. It is calculated by measuring the volume of $CO_2$ exhaled divided by the volume of $O_2$ consumed during respiration over time.
- RQ = 1.0 (Carbohydrates): Glucose oxidation uses 6 $O_2$ to produce 6 $CO_2$. ($6/6 = 1$).
- RQ = ~0.7 (Lipids): Because lipids have so little oxygen inherently, they require a massive intake of $O_2$ from the air to be fully oxidized into $CO_2$ and $H_2O$. Thus, the denominator ($O_2$ consumed) is much larger.
- RQ = ~0.9 (Proteins): An intermediate value.
Essential IMAT Glossary
📝 Mastery Practice: IMAT Biomolecules Quiz
Test your comprehensive understanding of the advanced carbohydrate and lipid biochemistry, molecular structures, and clinical lab analytics covered in this module. These questions are meticulously styled after the rigorous, integrated, multi-step logic required for the IMAT examination.