Meditaliano IMAT Prep
Session 19: Nervous System, Receptors, Effectors & Homeostasis
Introduction: The Symphony of Control and Response
Overview: The Human Nervous System

The Central Nervous System (CNS) processes information, while the Peripheral Nervous System (PNS) relays signals between the CNS and the rest of the body.
Nervous System Architecture & Information Flow

Nervous System Overview: This comprehensive diagram visualizes the CNS and PNS, including the detailed breakdown of the Autonomic system into Sympathetic and Parasympathetic pathways. At the bottom, the 5-step pathway maps the flow of information from Stimulus $\to$ Receptor $\to$ Sensory Neuron $\to$ CNS $\to$ Motor Neuron $\to$ Effector $\to$ Response.
Overview: The Sensory System

Receptors detect internal and external stimuli and transduce them into electrical impulses.
Welcome to one of the most heavily tested, high-yield sections of the IMAT Biology syllabus. Survival depends on an organism's ability to detect changes in its environment (stimuli), process this information, and execute an appropriate response. This rapid communication network is governed by the Nervous System, utilizing swift electrical impulses travelling up to 120 m/s. Simultaneously, long-term metabolic stability and homeostasis are maintained by the Endocrine System via chemical messengers in the blood.
This masterclass comprehensively details the intricate mechanisms of neural transmission, the anatomy and physiology of sensory organs (the eye and the ear), the molecular mechanics of skeletal muscle contraction, and the foundational principles of hormonal feedback loops.
IMAT Learning Objectives Checklist
- LO 19.1: Cell Biology of Neurons. Understand resting potentials, Nernst gradients, action potential phases (including refractory periods), and detailed synaptic transmission mechanisms.
- LO 19.2: Neuroanatomy & Organization. Differentiate CNS vs. PNS, identify brain lobes and spinal cord functions, and compare Sympathetic vs. Parasympathetic pathways.
- LO 19.3: Sensory Receptors. Master the optics of the eye, the phototransduction cascade in the retina, auditory mechanics in the cochlea, and vestibular equilibrium.
- LO 19.4: Effector Mechanisms. Deconstruct the sarcomere, fully explain the Sliding Filament Theory, differentiate muscle fiber types, and understand the energetics of contraction.
- LO 19.5: Endocrine Homeostasis. Compare peptide vs. steroid hormone actions, master the Hypothalamic-Pituitary axis, and map out negative feedback loops for blood glucose, temperature, and water balance.
Part 1: The Nervous System - Wiring the Body
1.1 Advanced Anatomy of Neurons & Glial Cells
The nervous tissue consists of two primary cell populations: Neurons (highly specialized, excitable cells that conduct electrical impulses) and Neuroglia (non-excitable supporting cells that outnumber neurons up to 10 to 1 and form the vital structural and metabolic matrix of the nervous system).
Visual: Simplified Neuron Structure

The unidirectional flow of information: Dendrite $\to$ Cell Body $\to$ Axon $\to$ Synapse.
The Physics of Saltatory Conduction
In unmyelinated fibers, an action potential must depolarize every adjacent microscopic segment of the axolemma (continuous conduction, ~1-2 m/s). In myelinated neurons, the lipid-rich myelin sheath acts as a high-resistance electrical insulator. Voltage-gated $Na^+$ and $K^+$ channels are concentrated exclusively at the unmyelinated gaps known as the Nodes of Ranvier.
The action potential cannot propagate through the myelin. Instead, the depolarizing current spreads rapidly through the axoplasm underneath the myelin and triggers the next node. The signal appears to "leap" from node to node. This increases transmission speeds dramatically (up to 120 m/s) and conserves massive amounts of cellular ATP, as the energy-hungry $Na^+/K^+$ pumps only need to actively restore gradients at the nodes rather than along the entire axon membrane.
Classification of Neurons
| Neuron Type | Functional Role | Structural Characteristics |
|---|---|---|
| Sensory (Afferent) Neuron | Transmits sensory data (light, touch, pain) from peripheral receptors towards the CNS. | Typically pseudo-unipolar. Features a single process that splits into a long peripheral axon (acting like a dendrite) and a central axon. The cell body is housed outside the CNS in a swelling called the Dorsal Root Ganglion. |
| Relay (Interneuron) | Integrates, processes, and routes information. The "calculators" of the brain. They form networks strictly within the CNS. | Multipolar. Highly branched, short dendrites and a short, often unmyelinated axon, optimizing them for local, rapid communication. |
| Motor (Efferent) Neuron | Carries executive motor commands away from the CNS to Effectors (skeletal muscles, smooth muscles, or glands). | Multipolar. Short dendrites and a single, very long, heavily myelinated axon that can extend over a meter. Cell body is located within the grey matter of the CNS. |
Deep Dive: Glial Cells (Neuroglia) - A Classic IMAT Trap

Notice how one oligodendrocyte grasps multiple axons, while one Schwann cell hugs only one axon segment.
Myelination is crucial for vertebrate nervous systems. The specific cells responsible differ based on location, which is a frequent subject of advanced biology questions:
- Oligodendrocytes (CNS): Found exclusively in the brain and spinal cord. A single oligodendrocyte extends multiple broad, flat processes to wrap around and myelinate multiple adjacent axons simultaneously. They are the target of autoimmune destruction in Multiple Sclerosis (MS).
- Schwann Cells (PNS): Found in peripheral nerves. A single Schwann cell wraps its entire cell body concentrically around a single segment of a single axon. Unlike oligodendrocytes, Schwann cells create a regeneration tube that actively assists in peripheral nerve regeneration after injury.
- Astrocytes (CNS): The most abundant glial cells. Star-shaped cells that regulate the chemical environment, mop up excess $K^+$ ions, recycle neurotransmitters (like glutamate to prevent excitotoxicity), and use their end-feet to wrap around capillaries, forming the highly restrictive Blood-Brain Barrier (BBB).
- Microglia (CNS): The resident immune cells. Originating from embryonic mesoderm (unlike other glia derived from ectoderm), they act as phagocytes/macrophages, wandering through brain tissue to clear cellular debris and defend against pathogens.
- Ependymal Cells (CNS): Ciliated epithelial cells that line the brain ventricles and central canal of the spinal cord, responsible for producing and circulating Cerebrospinal Fluid (CSF).
1.2 The Action Potential: Electrophysiology & Ion Dynamics
Neurons communicate via sudden, transient reversals of their membrane polarity. To understand the action potential, we must deeply understand the baseline state: the resting membrane potential.
Visual: The Electrogenic Sodium-Potassium Pump ($Na^+/K^+$ ATPase)

Primary Active Transport: The ubiquitous pump utilizes the hydrolysis of one ATP molecule to forcibly eject 3 $Na^+$ ions out of the cell and import 2 $K^+$ ions into the cell against their respective concentration gradients.
Establishing the Resting Membrane Potential (approx. -70 mV):
The interior of a resting neuron is negatively charged relative to the extracellular fluid. This polarized state is a delicate equilibrium maintained by three primary factors:
- Differential Permeability (Leak Channels): The membrane possesses non-gated "leak" channels for both sodium and potassium. Crucially, the membrane is approximately 50 to 100 times more permeable to $K^+$ than to $Na^+$. Therefore, positive $K^+$ ions constantly diffuse out of the cell down their steep concentration gradient, carrying positive charge away from the interior.
- The $Na^+/K^+$ Pump: Because it pumps 3 positive charges out for every 2 it brings in, it results in a net loss of one positive charge per cycle, directly contributing slightly to the negative resting potential (about -4 mV of the total -70 mV). Its primary job is maintaining the steep concentration gradients.
- Intracellular Anions: The cytoplasm is packed with large, negatively charged proteins, nucleic acids, and phosphate ions that cannot pass through the lipid bilayer, anchoring a negative charge inside the cell.
The Chronology of an Action Potential
An action potential is a rapid, all-or-nothing electrochemical explosion triggered by voltage-gated ion channels.
- Stimulus & Threshold: A graded potential slightly depolarizes the membrane. If this depolarization reaches the critical threshold potential of roughly -55 mV, a positive feedback loop is triggered. According to the All-or-None Law, crossing threshold guarantees a full, identical action potential. If it falls short (-56 mV), absolutely nothing happens.
- Rapid Depolarization (+40 mV): Reaching threshold instantly snaps open fast voltage-gated $Na^+$ channels. $Na^+$ experiences a massive electrochemical driving force and floods INTO the cell. The membrane potential reverses rapidly, skyrocketing past 0 mV to peak at roughly +35 mV to +40 mV.
- Repolarization: At the peak voltage, the fast $Na^+$ channels automatically close via a physical inactivation gate. Simultaneously, slower voltage-gated $K^+$ channels finally reach their fully open state. Driven by the new positive charge inside the cell and their concentration gradient, $K^+$ rapidly rushes OUT of the cell, dropping the membrane potential back toward negative resting values.
- Hyperpolarization (Undershoot): Voltage-gated $K^+$ channels are sluggish to close. Even as the membrane passes the -70 mV mark, $K^+$ continues to leak out, causing the membrane potential to dip below the normal resting level (plunging near the Nernst equilibrium potential for potassium, approx. -80 mV to -90 mV).
- Restoration: The voltage-gated channels reset to their closed states. The $Na^+/K^+$ pump and leak channels stabilize the membrane precisely back at the -70 mV resting potential, cocking the spring for the next firing.
High Yield Concept: Absolute vs. Relative Refractory Periods
Refractory periods are critical for two reasons: they limit the maximum firing frequency of a neuron, preventing seizures, and they force the action potential to travel unidirectionally down the axon.
- Absolute Refractory Period: Spans the entirety of the depolarization phase and the majority of the repolarization phase. The voltage-gated $Na^+$ channels are not just closed; they are physically blocked by their inactivation "plug".
Consequently, absolutely no stimulus, regardless of how intensely powerful it is, can trigger a second action potential. - Relative Refractory Period: Corresponds exactly with the hyperpolarization (undershoot) phase. The $Na^+$ channels have removed their inactivation plugs and returned to their standard "closed but ready" resting state. However, because the membrane is hyperpolarized (e.g., at -85 mV), it is much further away from the -55 mV threshold than normal. A new action potential can be fired, but it requires a significantly stronger-than-normal suprathreshold stimulus.
Visual: Action Potential Graph & Phases Detail

Advanced Electrophysiology & Ion Dynamics

Action Potential & Ion Dynamics: This comprehensive graph shows the membrane potential (mV) over time (ms), detailing the resting state (-70mV), threshold (-55mV), depolarization, repolarization, and hyperpolarization phases. Below the graph, 6 cross-sectional diagrams correspond to each phase, illustrating the specific states of Na+ and K+ channels and the resulting ion flow.

Advanced Schematic: A deeper look at the electrophysiological states and absolute vs. relative refractory periods, showing the physical state of the gated channels.
1.3 Synaptic Transmission: Chemical Communication
When an electrical action potential reaches the terminal arborizations (boutons) of an axon, it cannot jump across the empty space. The electrical signal must be converted into a chemical messenger to cross the synaptic cleft (a microscopic gap of roughly 20-30 nm) and bind to receptors on the target cell.
Architecture of a Chemical Synapse
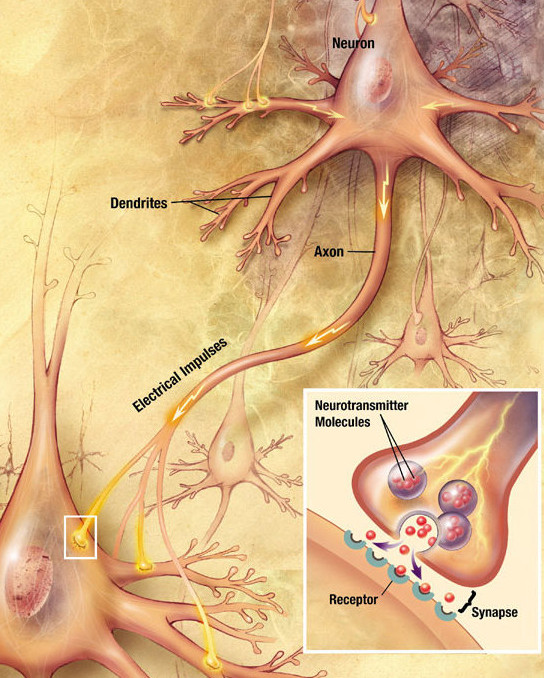
The Synaptic Transfer Cycle
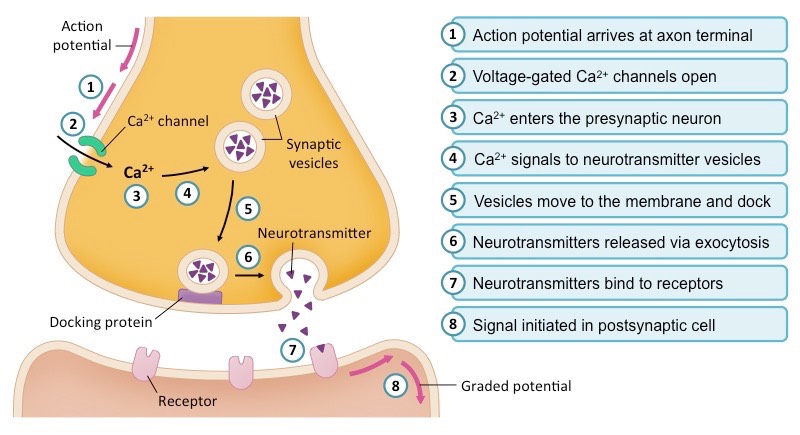
Molecular Synaptic Transmission

Synaptic Transmission: Details the presynaptic terminal, cleft, and postsynaptic membrane. Shows Ca2+ influx triggering 5 stages of neurotransmitter exocytosis. Compares EPSP (depolarization) and IPSP (hyperpolarization) mechanisms, explains spatial vs. temporal summation, and illustrates neurotransmitter termination.
Advanced Synaptic Integration

Signal Termination: Advanced schematic of synaptic integration and the mechanisms for clearing neurotransmitters (Enzymatic, Reuptake, Diffusion) from the synaptic cleft.
The Molecular Ballet of Exocytosis
-
1. Arrival & Calcium Influx:
The wave of depolarization from the action potential arrives at the presynaptic terminal, triggering the opening of specialized voltage-gated Calcium ($Ca^{2+}$) channels. Because extracellular calcium concentration is much higher, $Ca^{2+}$ ions rush rapidly into the presynaptic bouton.
-
2. Vesicle Docking and Exocytosis:
The sudden influx of $Ca^{2+}$ acts as an intracellular signaling ion. It binds to sensor proteins, causing structural changes in SNARE complexes. These complexes physically drag membrane-bound synaptic vesicles (packed with neurotransmitters) to the presynaptic membrane, forcing them to fuse. The neurotransmitters are violently dumped into the synaptic cleft via exocytosis.
-
3. Binding & Postsynaptic Potentials:
Neurotransmitters diffuse across the cleft and act as ligands, binding to specific ligand-gated ion channels on the postsynaptic membrane.
Excitatory (EPSP):Neurotransmitters (e.g., Glutamate) open $Na^+$ channels. The influx of positive $Na^+$ ions causes a localized depolarization, nudging the membrane closer to threshold.
Inhibitory (IPSP):Neurotransmitters (e.g., GABA) open $Cl^-$ channels (in) or $K^+$ channels (out). Both cause a localized hyperpolarization, silencing the neuron.
Neural Integration (Summation): The Biological Calculator
A single postsynaptic neuron receives synapses from up to 10,000 different presynaptic neurons simultaneously. The Axon Hillock continuously sums these inputs mathematically. If the net sum of all depolarizing EPSPs outweighs the hyperpolarizing IPSPs and pushes the voltage past -55mV, the neuron fires.
Multiple different presynaptic neurons fire simultaneously at different physical spots. Their individual EPSPs pool together geographically.
A single presynaptic neuron fires repetitively and rapidly. The second EPSP arrives before the first repolarizes, stacking on top of each other over time.
Crucial Step: The Termination of the Signal (Neurotransmitter Clearance)
For a synapse to function as a precise communication tool rather than causing chaotic, sustained tetanus, the neurotransmitter must be rapidly cleared from the cleft. The nervous system employs three distinct mechanisms:
-
Enzymatic Degradation: Destructive enzymes anchored in the cleft cleave the neurotransmitter.
Classic Example: Acetylcholinesterase (AChE) at the neuromuscular junction hydrolyzes ACh into acetate and choline. Nerve gases (like Sarin) inhibit AChE, leading to fatal, continuous muscle convulsions. -
Active Reuptake: Transport proteins on the presynaptic membrane actively pump the intact neurotransmitter back into the terminal for repackaging.
Clinical Relevance: SSRIs (antidepressants like Prozac) block serotonin reuptake pumps, artificially leaving serotonin in the cleft longer. - Passive Diffusion & Glial Clearance: Neurotransmitters diffuse out of the cleft. Nearby astrocytes frequently assist by actively absorbing escaping neurotransmitters (particularly glutamate) to prevent neurotoxic build-up.
1.4 Central & Peripheral Nervous System Organization
The gross anatomical structure of the human nervous system is rigidly divided into the Central Nervous System (CNS) and the Peripheral Nervous System (PNS).
Central Nervous System (CNS)
Comprises the Brain and Spinal Cord. The integration and command center.
- Cerebrum (Cortex): Divided into 4 lobes.
- Frontal: Executive function, personality, primary motor cortex (precentral gyrus).
- Parietal: Primary somatosensory cortex (postcentral gyrus - touch, pain).
- Temporal: Auditory processing, language comprehension (Wernicke's), memory (Hippocampus).
- Occipital: Visual processing center.
- Cerebellum: "Little brain." Crucial for motor coordination, posture, and learning fine motor skills (muscle memory). It acts as an error-correction machine.
- Diencephalon: Contains the Thalamus (sensory relay station) and Hypothalamus (master regulator of homeostasis and endocrine system).
- Brainstem: Midbrain, Pons, and Medulla Oblongata. Controls vital autonomic, life-sustaining functions (respiratory rhythm, heart rate, blood pressure).
- Spinal Cord: Uniquely, its Grey Matter is on the inside (butterfly shape), and White Matter (myelinated tracts) is on the outside.
Peripheral Nervous System (PNS)
Consists of 12 pairs of cranial nerves and 31 pairs of spinal nerves. Divided into Somatic (voluntary) and Autonomic (involuntary) branches.
The Autonomic Nervous System (ANS)
Maintains internal balance using two mutually antagonistic branches that dual-innervate most organs.
| Physiological Target | Sympathetic ("Fight or Flight") |
Parasympathetic ("Rest & Digest") |
|---|---|---|
| Pupils | Dilate (Mydriasis) | Constrict (Miosis) |
| Heart Rate | Accelerates & stronger | Decelerates (Vagus nerve) |
| Airways | Bronchodilation | Bronchoconstriction |
| Digestion | Inhibited (blood diverted) | Stimulated (peristalsis up) |
| Liver | Glycogenolysis (release glucose) | Glycogenesis (store glucose) |
| Neurotransmitter at Effector | Noradrenaline (Norepinephrine) | Acetylcholine (ACh) |
Clinical Definition: Brain Death vs. Vegetative State
Understanding the functional hierarchy of the brain is crucial for medical ethics.
Irreversible cessation of all brain function, including the cerebrum AND the vital Brainstem. Because the medulla oblongata is dead, the patient cannot breathe spontaneously. Legally and physiologically deceased. Reflexes mediated by cranial nerves are absent.
Severe trauma has destroyed the Cerebrum (loss of consciousness/personality), but the primitive Brainstem remains intact. The patient breathes spontaneously, maintains a heartbeat, and regulates blood pressure, but there is "nobody home."
1.5 The Spinal Cord & Reflex Arcs: Hardwired Survival Machinery
A reflex is an involuntary, rapid, and unlearned motor response to a specific stimulus. It exists to protect the body from acute damage by bypassing slow conscious processing in the higher brain. The neural circuitry driving a reflex is called a Reflex Arc.
The Bell-Magendie Law (Fundamental Neuroanatomy)
A fundamental rule governing how spinal nerves connect to the cord. A spinal nerve is "mixed" (carrying both sensory and motor fibers). Just before connecting to the spinal cord, it splits:
Strictly Sensory (Afferent). Information *enters* the spinal cord via the back door. Houses the Dorsal Root Ganglion.
Strictly Motor (Efferent). Commands *exit* the spinal cord via the front door.
Mnemonic: SAME DAVE
Sensory Afferent, Motor Efferent. | Dorsal Afferent, Ventral Efferent.
Visual: Anatomy of a Spinal Reflex & The 5-Part Pathway

The standard 5-part pathway:
1. Receptor (detects stimulus) $\to$ 2. Sensory Neuron (travels via dorsal root) $\to$ 3. Integration Center (Grey matter of spinal cord) $\to$ 4. Motor Neuron (exits via ventral root) $\to$ 5. Effector (muscle contracts).
Functional Reflex Arc Anatomy & Spinal Organization

Spinal Organization: This diagram highlights the anatomy of a reflex arc, showing the sensory input through the dorsal root and motor output through the ventral root, perfectly illustrating the Bell-Magendie Law.
Monosynaptic Reflex (Stretch Reflex)
The fastest, simplest reflex arc. There is only ONE central synapse. The sensory neuron synapses directly onto the motor neuron inside the spinal cord. NO interneurons.
Classic Example: Patellar Reflex (Knee-Jerk)
- Tapping the patellar ligament stretches the quadriceps muscle.
- Stretch receptors (muscle spindles) fire an action potential.
- The sensory neuron enters the spinal cord and directly excites the motor neuron for the same quadriceps muscle.
- The quadriceps contracts instantly, kicking the leg.
Polysynaptic Reflex (Flexor Withdrawal)
A more complex reflex designed to pull limbs rapidly away from painful stimuli. It involves one or more Interneurons (relay neurons), meaning there are at least two synapses.
Classic Example: Touching a Hot Stove
- Nociceptors (pain receptors) detect extreme heat.
- Sensory neuron fire and enters the dorsal horn.
- It synapses onto several Interneurons, which perform complex routing (Reciprocal Inhibition):
- EXCITE: Excitatory signal to flexor muscles (biceps) to contract and yank the arm away.
- INHIBIT: Inhibitory signal to antagonistic extensor muscles (triceps) to relax so they don't fight the movement.
Part 2: Sensory Physiology - Vision and The Eye
The eye is an exquisite optical instrument engineered to capture photons of light, precisely focus them onto a cellular canvas, and transduce that electromagnetic energy into a massive stream of action potentials.
2.1 Advanced Anatomy of the Optical Pathway
Internal Tissues and Macula
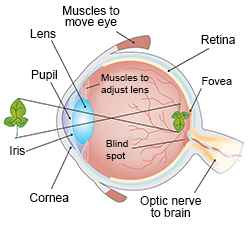
The Path of Light: From Air to Retina
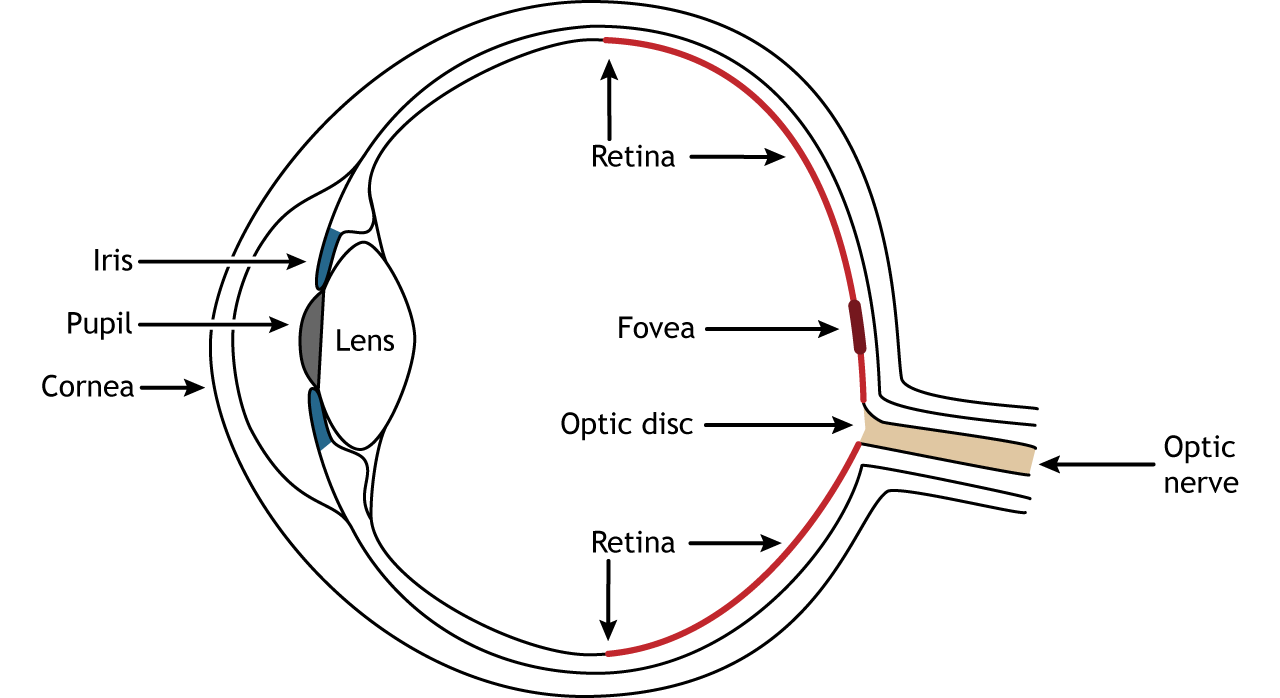
- Cornea: Light first strikes this transparent, dome-shaped window (a continuation of the tough white Sclera). It performs ~70-80% of the gross refractive bending of light. It is avascular.
- Aqueous Humor & Pupil: Light passes through the watery anterior chamber, then through the Pupil (the variable aperture in the center of the colored Iris).
- The Lens: A flexible, crystalline, biconvex structure. While the cornea does heavy bending, the lens fine-tunes the refraction to achieve a razor-sharp focal point.
- Vitreous Humor: Light traverses the large posterior chamber filled with a clear gelatinous mass that maintains eyeball shape and presses the retina flat.
- Retina: The innermost sensory layer where phototransduction occurs. Contains the Macula / Fovea Centralis (packed exclusively with cones for highest visual acuity) and the Optic Disc (the blind spot where ganglion cell axons exit to form the Optic Nerve).
2.2 Optics and Muscular Control: Accommodation
To shift our focus dynamically from a distant mountain to a book held close, the elastic crystalline Lens must change its curvature. This is driven by the smooth muscles of the Ciliary Body, connected to the lens by Suspensory Ligaments.
Ocular Optics & Muscular Control: This diagram illustrates the biomechanics of accommodation, showing how the ciliary muscle's contraction/relaxation alters the tension on the suspensory ligaments to change the lens shape for near vs. distant vision.
| Visual Target | Incoming Light | Ciliary Muscle Action | Suspensory Ligaments | Resulting Lens Shape | Refractive Power |
|---|---|---|---|---|---|
| Distant (> 6 meters) |
Almost parallel | Relax (ring expands) | Pulled Taut (Tight) | Flattened (Thin) | Low |
| Near (Reading) |
Highly divergent | Contract (moves inward) | Go Slack (Loose) | Spherical (Thick/Round) | High |
Refractive Errors (Defects in Optics)
Eyeball is too long, or cornea bends light too sharply. Light from distant objects focuses in front of the retina.
Correction: Diverging (Concave) lens.
Eyeball is too short, or lens is weak. Light from near objects fails to converge quickly enough, focusing behind the retina.
Correction: Converging (Convex) lens.
As humans age, crystalline proteins harden and lose natural elasticity. When ciliary muscles contract for near vision, the stiff lens physically cannot spring back into a round shape. Near-point focusing fails.
2.3 The Pupillary Light Reflex
An autonomic reflex to protect the retina from phototoxicity while maximizing vision in dim light. The iris contains an antagonistic pair of smooth muscles.
| Environment | Autonomic Branch | Active Iris Muscle | Pupil State |
|---|---|---|---|
| Bright Sunlight | Parasympathetic (Cranial Nerve III) |
Circular Sphincter contracts | Constricts (Miosis) |
| Dim Starlight | Sympathetic | Radial Dilator contracts | Dilates (Mydriasis) |
2.4 The Retina: Photoreceptors & The Inverted Architecture
Visual: The "Inverted" Layers of the Retina
The Evolutionary Oddity: Counterintuitively, light-sensitive photoreceptors are buried at the very back. Incoming light must pass through ganglion cell axons, ganglion cell bodies, amacrine cells, and bipolar cells before it hits the rods and cones. The electrical signal then travels in the reverse direction: Photoreceptors $\to$ Bipolar Cells $\to$ Ganglion Cells $\to$ Optic Nerve.
Retina Architecture & Photoreceptor Function

Visual Mastery: The left panel illustrates the inverted structure of the retina. The right panel provides a side-by-side comparison of Rods and Cones (Duplicity Theory), and the bottom right details the molecular mechanics of the 'Dark Current' vs. 'Light' activation.
The Duplicity Theory of Vision: Rods vs. Cones
Human vision relies on two completely distinct functional systems operating in parallel.
| Feature |
Rods
(~120 million per eye)
|
Cones
(~6 million per eye)
|
|---|---|---|
| Function | Scotopic Vision (Night). Monochromatic (Black & White). Detects motion and shapes. | Photopic Vision (Day). Color vision. High-resolution fine detail. |
| Photopigment | Rhodopsin (Visual Purple). Bleaches instantly in bright light. Takes ~30-45 mins to regenerate in the dark. | Photopsins (Iodopsins). Three distinct types responding to Red, Green, and Blue wavelengths for trichromatic vision. |
| Light Sensitivity | Extremely High. Can respond to a single photon. | Low. Requires massive barrages of thousands of photons to activate. |
| Visual Acuity (Resolution) | Very Low (Blurry). High Convergence. Up to 100 rods connect to a single bipolar cell (high spatial summation). | Extremely High (Sharp). Zero convergence in the fovea (1 cone $\to$ 1 bipolar cell $\to$ 1 ganglion cell) giving pinpoint 4K resolution. |
| Distribution | Dense in the peripheral retina. Completely absent from the fovea. | Highly concentrated in the Fovea Centralis. Drops off precipitously in the periphery. |
The Molecular Masterpiece: Phototransduction and the "Dark Current"
Sensory neurons usually sit at a negative resting potential and fire when stimulated. Photoreceptors do the exact opposite. They are highly counterintuitive.
-
In the Dark1. The State of Darkness (The Dark Current)
In a pitch-black room, rod cells are highly active. High intracellular levels of cyclic GMP (cGMP) hold thousands of ligand-gated $Na^+$ channels continuously open. Sodium constantly floods into the cell (the "Dark Current").
This keeps the rod membrane constantly depolarized (around -40 mV). Because it is depolarized, the rod continuously releases a steady stream of the inhibitory neurotransmitter Glutamate from its synaptic terminal. This glutamate constantly inhibits the downstream Bipolar Cell, preventing it from firing. The brain receives no signal, and perceives "darkness."
-
In the Light2. The Flash of Light (The Biochemical Cascade)
When a single photon strikes the rod:
- The photon is absorbed by Rhodopsin. Specifically, it hits the chromophore 11-cis-retinal.
- The energy forces the 11-cis-retinal to instantly snap into a straight all-trans-retinal shape ("bleaching").
- This shape change activates the opsin protein, which activates a G-protein called Transducin.
- Transducin activates an enzyme called Phosphodiesterase (PDE).
- PDE rapidly destroys the cGMP molecules in the cytoplasm.
- With cGMP destroyed, the ligand-gated $Na^+$ channels snap CLOSED.
- The Dark Current is severed. The rod cell membrane rapidly plummets to -70 mV, becoming HYPERPOLARIZED.
- Crucially, because the rod is hyperpolarized, voltage-gated calcium channels close, and the rod STOPS releasing the inhibitory Glutamate.
- Released from its constant inhibition, the downstream Bipolar Cell instantly depolarizes, firing an action potential to the Ganglion cell, which fires a spike train down the optic nerve. The brain registers light.
Visual: Neural Pathways of Vision & The Optic Chiasm
The Golden Rule of Visual Pathways: Only the fibers from the medial (nasal) half of each retina cross over at the Optic Chiasm. Fibers from the lateral (temporal) half stay on the same side. This ensures that the right hemisphere of the brain processes the entire left visual field, and vice versa.
Part 3: Sensory Physiology - The Ear & Equilibrium
The human ear is a dual-function biomechanical marvel housing the intricate labyrinths responsible for audition (hearing) and the vestibular system (equilibrium and spatial orientation).
3.1 Gross Anatomy of the Ear
Visual: Cross-Section Anatomy of the Human Ear

The Outer Ear
Consists of the Pinna (Auricle), which funnels sound waves down the auditory canal, terminating at the Tympanic Membrane (Eardrum). Airborne sound waves strike the eardrum, causing it to vibrate with exact matching frequency and amplitude.
The Middle Ear
A small cavity containing the Ossicles: Malleus, Incus, and Stapes. The Stapes plunges into the Oval Window.
The Eustachian Tube connects to the throat to equalize air pressure, preventing the eardrum from rupturing.
The Inner Ear
An incredibly complex, fluid-filled labyrinth of bone and membrane. It contains the snail-shell shaped Cochlea (dedicated to hearing) and the Vestibular Apparatus (dedicated to balance).
Advanced Biophysics: Impedance Matching in the Middle Ear
The fundamental challenge of hearing: Sound waves travel easily through thin air, but the sensory hair cells are submerged in the dense fluid of the inner ear. If airborne sound struck the fluid directly, over 99% of the sound energy would reflect off the surface. You would be deaf.
The Middle Ear solves this by acting as an acoustic transformer, amplifying the pressure by roughly 22 times:
- The Area Ratio: The massive Tympanic Membrane gathers acoustic force over a huge area and concentrates it onto the microscopic footprint of the Stapes on the Oval Window. ($Pressure = Force / Area$). This provides ~17x amplification.
- The Lever Action: The ossicular bones are articulated as an asymmetrical lever system, multiplying the force by ~1.3x.
$17 \times 1.3 \approx 22.1$ times amplification
3.2 The Mechanics of Hearing: The Cochlea and Tonotopy
Visual: Cross-Section of the Cochlea & The Organ of Corti
The biological microphone: The Organ of Corti rests upon the flexible Basilar Membrane. When fluid waves ripple the membrane, the stereocilia of the hair cells are sheared against the rigid overlying Tectorial Membrane.
The Detailed Pathway of Transduction
- The stapes pumps against the oval window, creating hydraulic pressure waves in the perilymph fluid of the cochlea.
- These waves press down on the cochlear duct, causing the rubbery Basilar Membrane to ripple up and down.
- Sitting atop this bouncing membrane is the Organ of Corti, containing mechanoreceptor Hair Cells.
- The stiff microvilli (Stereocilia) of the hair cells are embedded in the rigid Tectorial Membrane above.
- As the basilar membrane bounces but the tectorial membrane stays still, a shearing force physically bends the stereocilia.
- The Molecular Trigger: Bending the hairs pulls open mechanically-gated ion channels via microscopic "tip links."
- Because endolymph fluid is uniquely rich in Potassium ($K^+$), $K^+$ rapidly floods into the hair cell (opposite of normal neurons), depolarizing it.
- The depolarized cell opens voltage-gated calcium channels, releasing Glutamate to excite the auditory nerve.
Tonotopy: The Frequency Map of the Basilar Membrane
The cochlea physically acts as an acoustic prism, spatially mapping pitch (frequency) along the length of the basilar membrane.
The basilar membrane is narrow, thick, and stiff. It requires tremendous energy to move. Only very fast, high-energy waves (High Frequency / High Pitch, ~20,000 Hz) create a resonance wave here.
The basilar membrane is wide, thin, and floppy. Slow, long-wavelength sounds travel all the way up the spiral and create a maximum amplitude wave here (Low Frequency / Low Pitch, ~20 Hz).
3.3 The Vestibular System (Equilibrium & Balance)
Located adjacent to the cochlea, this system detects head movement, rotation, and gravity to maintain balance and coordinate eye movements.
Semicircular Canals (Dynamic Equilibrium)
Three fluid-filled tubes arranged orthogonally in the X, Y, and Z planes. They detect Rotational Acceleration. When you rotate your head (e.g., spinning or shaking your head "no"), the dense fluid inside lags behind due to inertia. This moving fluid physically bends a gelatinous cupula, which bends the hair cells inside, firing action potentials.
Otolith Organs (Static Equilibrium)
The Utricle and Saccule detect the position of your head relative to gravity (tilting) and Linear Acceleration (riding in an elevator or accelerating in a car). They contain tiny, heavy calcium carbonate crystals (Otoliths) resting on a jelly layer over hair cells. Gravity pulls these heavy crystals downward when you tilt your head, bending the hairs.
Part 4: Effectors - Skeletal Muscle Mechanics
Effectors execute the commands of the nervous system. Skeletal muscle is striated, multinucleated, and under voluntary (somatic) control. It converts chemical energy (ATP) directly into mechanical force.
4.1 Microscopic Anatomy: The Sarcomere
A skeletal muscle fiber (cell) is packed with hundreds of rod-like Myofibrils. Myofibrils consist of repeating contractile units called Sarcomeres, formed by overlapping arrays of protein myofilaments: Thick Myosin and Thin Actin.
Visual: Sarcomere Banding Pattern Detail

Visual: Sliding Filament Mechanics & Cellular Energetics

Sliding Filament Model: This diagram illustrates the molecular interaction between actin and myosin, the role of Calcium in exposing binding sites, and the ATP-driven power stroke cycle.
Decoding the Sarcomere Bands (Highly Tested IMAT Concept)
- Z-line (disc): The zig-zag boundary of a single sarcomere. Thin actin filaments are rigidly anchored here.
- A-band: The dark band. Represents the entire length of the thick Myosin filaments.
Crucial Fact: The A-band NEVER changes length during a contraction. - I-band: The light band. Contains only thin Actin filaments (no myosin overlap).
Changes: This band shortens and can disappear during contraction as actin slides inward. - H-zone: The center of the A-band containing only Myosin (no actin overlap).
Changes: This zone shrinks and completely disappears during a maximal contraction. - M-line: The structural midline of the sarcomere holding myosin in place.
4.2 The Sliding Filament Theory
Muscle contraction occurs when actin and myosin filaments slide past each other, shortening the sarcomere without the filaments themselves changing length. This process relies on Excitation-Contraction Coupling, requiring both Calcium and ATP.
- Action Potential Propagation: The motor end plate depolarizes, and the action potential sweeps across the sarcolemma, diving deep into the core of the muscle fiber via hollow invaginations called T-tubules.
- Calcium Release: The voltage change in the T-tubule triggers the adjacent Sarcoplasmic Reticulum (SR) to throw open massive calcium release channels. $Ca^{2+}$ floods the sarcoplasm.
- Uncovering the Binding Site: Normally, the regulatory protein Tropomyosin blocks the myosin-binding sites on actin. When $Ca^{2+}$ binds to Troponin, it causes a conformational change that yanks tropomyosin away, exposing the binding sites.
- Cross-Bridge Formation: Myosin heads (pre-cocked into a high-energy state by hydrolyzing ATP $\to$ ADP + Pi) immediately grab the exposed actin sites, forming a cross-bridge.
- The Power Stroke: The myosin head releases the Pi and ADP. This release of energy causes the myosin head to pivot forcefully, ratcheting the actin filament toward the M-line. The sarcomere shortens.
- Detachment: A fresh molecule of ATP binds to the myosin head. This breaks the actin-myosin bond, allowing detachment.
Clinical Correlate: In Rigor Mortis, the body runs out of ATP after death. Myosin cannot detach from actin, leaving muscles locked in a state of permanent contraction. - Reset: The myosin head acts as an ATPase, hydrolyzing the new ATP to recock its head. Relaxation occurs when the SR uses active transport pumps to suck the $Ca^{2+}$ back into storage, allowing tropomyosin to slide back over the binding sites.
4.3 Energy Systems and Muscle Fiber Types
A contracting muscle requires immense amounts of ATP for millions of simultaneous myosin power strokes and active $Ca^{2+}$ pumping. It utilizes three sequential energy systems depending on the duration and intensity of exercise.
- Phosphagen System (Direct Phosphorylation): For immediate, explosive energy (0-10 seconds). Creatine Phosphate donates a placeholder group directly to ADP to instantly regenerate ATP. Does not require oxygen.
- Anaerobic Glycolysis: For high-intensity, short-duration work (10 seconds to 2 minutes). Breaks down glucose without oxygen, yielding a net of 2 ATP and Lactic Acid (which causes muscle burn and eventually fatigue).
- Aerobic Respiration (Oxidative Phosphorylation): For prolonged, endurance exercise. Uses oxygen in the mitochondria to completely break down glucose, glycogen, and fatty acids, yielding ~36 ATP per glucose. Slow to initiate but highly efficient and sustainable.
| Feature | Type I Fibers Slow Twitch / Slow Oxidative |
Type II Fibers Fast Twitch / Fast Glycolytic |
|---|---|---|
| Contraction Speed & Fatigue | Slow contraction. Highly resistant to fatigue. Can contract for hours. | Fast, powerful contraction. Fatigues very rapidly due to lactic acid build-up. |
| Primary Metabolism | Aerobic Respiration. | Anaerobic Glycolysis. |
| Cellular Features | Packed with Mitochondria. Dense capillary network. High levels of Myoglobin (an oxygen-storing protein), giving the muscle a dark RED color. | Few Mitochondria. Large glycogen stores. Low Myoglobin, giving the muscle a pale/WHITE color. |
| Typical Function | Maintaining postural tone, endurance running (Marathon). (e.g., Soleus muscle). | Explosive movements, sprinting, heavy weightlifting, jumping. (e.g., Gastrocnemius, eye muscles). |
Part 5: Endocrine System & Homeostasis
Homeostasis is the maintenance of a relatively constant internal environment (temperature, pH, blood glucose, osmolarity, calcium levels) despite massive external changes. While the nervous system handles rapid, targeted responses, the Endocrine System utilizes hormones circulating in the bloodstream to enact widespread, long-term metabolic changes, primarily regulated by Negative Feedback Loops.
Overview: Endocrine System & Major Feedback Loops

Endocrine Mastery Map: This diagram maps the major endocrine glands and details three vital negative feedback loops: Blood Glucose (Insulin/Glucagon), Thermoregulation, and Water Balance (ADH). It also contrasts Peptide vs. Steroid hormone mechanisms.
5.1 Nervous System vs. Endocrine System
| Feature | Nervous System | Endocrine System |
|---|---|---|
| Signal Type | Electrical (Action Potentials) & Chemical (Neurotransmitters crossing a micro-gap) | Chemical only (Hormones secreted directly into the systemic bloodstream) |
| Speed of Action | Extremely Fast (milliseconds) | Slow (minutes, hours, days, or months) |
| Duration of Effect | Short-lived (transient, terminates instantly when stimulus stops) | Long-lasting (can persist long after the initial stimulus is gone) |
| Target Area | Highly localized and specific (e.g., a single muscle fiber or gland) | Widespread (affects any cell in the entire body possessing the specific hormone receptor) |
5.2 Molecular Mechanisms of Hormones: Peptide vs. Steroid
Hormones only affect target cells that express the correct receptor. How a hormone physically interacts with that cell depends entirely on its biochemical structure.
Peptide/Protein/Amine Hormones
- Synthesized from amino acids. They are hydrophilic (water-loving).
- Because they are hydrophilic, they cannot cross the hydrophobic lipid bilayer of the cell membrane.
- Mechanism: They bind to extracellular receptors on the membrane surface. This activates a G-protein inside the membrane, which activates an enzyme (like Adenylate Cyclase) to produce a Second Messenger inside the cell (usually cyclic AMP, or cAMP). The second messenger then triggers a massive cascade of protein kinases that alter cellular activity (phosphorylation).
- Speed: Very fast acting because they activate enzymes already present in the cell.
- Examples: Insulin, Glucagon, Adrenaline (Epinephrine), ADH, Growth Hormone.
Steroid Hormones
- Synthesized from cholesterol. They are hydrophobic (lipid-loving).
- Because they are hydrophobic, they easily diffuse directly through the lipid bilayer into the interior of the cell.
- Mechanism: They bind to intracellular receptors located in the cytoplasm or inside the nucleus. The hormone-receptor complex then acts directly as a Transcription Factor, binding to specific regions of DNA to turn specific genes on or off, directing the synthesis of brand new proteins.
- Speed: Slower acting because it takes time to transcribe mRNA and translate new proteins, but effects are long-lasting.
- Examples: Testosterone, Estrogen, Progesterone, Cortisol, Aldosterone.
5.3 The Hypothalamic-Pituitary Axis (The Master Regulators)
The Hypothalamus acts as the executive bridge between the nervous system (detecting environmental shifts) and the endocrine system (ordering physiological responses). It dictates orders to the Pituitary gland (Hypophysis) situated just below it.
Visual: Major Pituitary Hormones

Posterior Pituitary (Neurohypophysis)
Not a true gland, but an extension of neural tissue from the hypothalamus. It does NOT synthesize its own hormones. It simply stores and releases two hormones produced by the hypothalamus via neurosecretory cells:
- Oxytocin: Stimulates uterine contractions during childbirth and milk ejection from mammary glands (One of the rare examples of a Positive Feedback loop).
- ADH (Antidiuretic Hormone / Vasopressin): Promotes water retention in the kidneys by inserting aquaporins into the collecting ducts, concentrating urine.
Anterior Pituitary (Adenohypophysis)
True glandular tissue. The hypothalamus secretes Releasing or Inhibiting hormones (e.g., TRH, CRH) into a specialized capillary network (hypophyseal portal system). These trigger the anterior pituitary to manufacture and release its own trophic hormones:
- TSH (Thyroid Stimulating Hormone $\to$ Thyroid $\to$ Metabolism)
- ACTH (Adrenocorticotropic Hormone $\to$ Adrenal Cortex $\to$ Cortisol for long-term stress)
- FSH & LH (Gonadotropins $\to$ Ovaries/Testes $\to$ Repro)
- GH (Growth Hormone $\to$ Bones/Muscles)
- Prolactin (Mammary glands $\to$ Milk production)
5.4 Key Homeostatic Loops
1. Thermoregulation (Hypothalamus)
The hypothalamus acts as the body's thermostat, adjusting mechanisms to keep core temp around 37°C.
- Vasodilation: Skin arterioles dilate (widen), allowing warm blood to flow near the skin surface to radiate heat away.
- Sweating: Glands secrete water for evaporative cooling.
- Behavioral: Lethargy, seeking shade.
- Vasoconstriction: Skin arterioles constrict (narrow), redirecting blood deep to the core to conserve vital heat.
- Shivering: Involuntary skeletal muscle contraction generates metabolic heat.
- Piloerection: Goosebumps trap an insulating air layer.
2. Blood Glucose Regulation (Pancreas)
Managed by the Islets of Langerhans in the Pancreas to keep glucose ~90mg/100ml.
Detected by Beta ($\beta$) cells. They secrete Insulin. Insulin causes body cells to take up glucose. It forces the Liver to convert soluble glucose into insoluble Glycogen for storage (Glycogenesis). Blood sugar falls.
Detected by Alpha ($\alpha$) cells. They secrete Glucagon. Glucagon targets the Liver, forcing it to break down stored Glycogen back into free glucose (Glycogenolysis) and release it. Blood sugar rises.
3. Osmoregulation / Water Balance (Kidneys & Brain)
Regulating the water potential of the blood prevents cells from bursting (lysis) or shrinking (crenation).
Hypothalamus detects concentrated blood. Posterior pituitary releases ADH. ADH inserts aquaporins into kidney Collecting Ducts, making them highly permeable to water. Water is reabsorbed into blood. Urine volume drops and is highly concentrated.
Hypothalamus detects dilute blood. ADH release is inhibited. Without ADH, collecting ducts become impermeable to water. Water remains in the filtrate. Urine volume increases massively and becomes pale and dilute.
Comprehensive Mastery Quiz (30 Questions)
Test your deep understanding of the advanced mechanisms covering neurobiology, sensory physiology, muscle mechanics, and endocrinology. These questions mimic the difficulty and analytical depth expected on the IMAT.