Lesson 7: Advanced Cell Mechanics, History & Diversity
A complete integration of advanced cellular physiology and comprehensive histology. From molecular transport to tissue architecture.
Introduction & Scope
This lesson bridges the gap between molecular cell biology and the structural organization of tissues (Histology). We will explore the history of cell discovery, the physics of membrane transport, signal transduction pathways, and the four fundamental tissue types found in the human body.
Part 1: Advanced Cell Biology
- History: From Hooke to Virchow.
- Membrane: Asymmetry, Lipid Rafts, Glycocalyx.
- Transport: Pumps, Channels, Cotransport thermodynamics.
- Signaling: GPCRs, Second Messengers, RTKs.
- Sorting: Signal sequences, Vesicular traffic.
Part 2: Histology (The 4 Tissues)
- Epithelium: Lining, Glands, Junctions.
- Connective: Bone, Blood, Cartilage, Proper.
- Muscle: Sarcomere dynamics, Calcium control.
- Nervous: Action Potential, Synapse, Glia.
Advanced Cell Biology
1.1 History of Cell Theory & Scale
The cell theory is the foundation of modern biology. It emerged from the work of several pioneers over two centuries.
Image 1: Timeline of Cell Theory and Biological Scales

Visual Analysis: History & Scale
Cell Theory Timeline (Left)
- 1665 (Robert Hooke): Observation of cork cells; coined the term "Cell".
- 1674 (Leeuwenhoek): Discovery of living "animalcules" (bacteria, etc.).
- 1831 (Robert Brown): Description of the cell nucleus.
- 1838/39 (Schleiden & Schwann): Establishment of the foundational Cell Theory.
- 1855 (Virchow): Addition of the third principle: "Omnis cellula e cellula" (All cells from cells).
Biological Scale (Right)
A comparison of resolution across three levels of observation:
- Human Vision: Resolves items like human hair.
- Light Microscope: Resolves RBCs, bacteria, and large viruses.
- Electron Microscope: Resolves proteins and individual atoms, highlighting the microscopic nature of cellular life.
| Year | Scientist | Discovery & Significance |
|---|---|---|
| 1665 | Robert Hooke | Used a primitive compound microscope to observe cork. Coined the term "Cell" (Latin cella: small room). Published Micrographia. |
| 1674 | Anton van Leeuwenhoek | "Father of Microbiology". Used high-quality single lenses to observe living cells (bacteria, sperm, protozoa), calling them "animalcules". |
| 1831 | Robert Brown | Described the Nucleus as a constant component of plant cells (observed in orchids). |
| 1838 | Matthias Schleiden | Stated that all plants are composed of cells. |
| 1839 | Theodor Schwann | Stated that all animals are composed of cells. Together with Schleiden, established the Cell Theory. |
| 1855 | Rudolf Virchow | Added the third tenet: "Omnis cellula e cellula" (All cells arise from pre-existing cells), refuting spontaneous generation. |
1.2 Membrane Architecture & Asymmetry
The plasma membrane is a dynamic Fluid Mosaic. Beyond the basic bilayer, it exhibits strictly regulated asymmetry crucial for cell function and survival.
Lipid Asymmetry
Phospholipids are synthesized on the cytosolic face of the Smooth ER. This creates an imbalance.
- Scramblases (ER): Randomly flip lipids to the luminal side to equalize layer growth. No ATP required.
- Flippases (Golgi/Plasma Membrane): Specific enzymes (P-type ATPases) that use ATP to move specific lipids (Phosphatidylserine, Phosphatidylethanolamine) to the cytosolic leaflet.
- Result: The outer leaflet is rich in Phosphatidylcholine and Sphingomyelin. The inner leaflet is rich in Phosphatidylserine (negative charge).
During apoptosis, enzymes called scramblases are activated (and flippases inhibited), exposing Phosphatidylserine on the outer surface. This serves as an "Eat Me" signal for macrophages.
Image 2: Dynamic Membrane Asymmetry and Apoptosis

Visual Analysis: Dynamic Membrane Asymmetry & Apoptosis
Generating Asymmetry (Left)
- Smooth ER: Site of lipid synthesis. Scramblases randomly flip lipids to equalize both leaflets (ATP-independent).
- Golgi/Plasma Membrane: Flippases (P-type ATPases) selectively move Phosphatidylserine (PS) and Phosphatidylethanolamine (PE) to the cytosolic leaflet (ATP-dependent).
- Final Distribution: Outer leaflet is rich in Phosphatidylcholine and Sphingomyelin; Inner leaflet is rich in PS (negative charge) and PE.
Asymmetry & Apoptosis (Right)
In healthy cells, flippases keep PS on the inside. In apoptotic cells:
- Flippase Inactivation: Stops the internal sequestration of PS.
- Scramblase Activation: Randomizes lipid distribution.
- "Eat Me" Signal: PS is exposed on the outer surface, signaling macrophages to engulf the cell.
1.3 Advanced Transport Mechanisms
Cells maintain distinct internal environments via selective transport. Transport is classified by energy requirement and direction relative to the electrochemical gradient.
| Type | Mechanism | Energy | Examples |
|---|---|---|---|
| Simple Diffusion | Directly through bilayer | Thermal (Entropy) | $O_2, CO_2, N_2$, Steroids, Ethanol |
| Facilitated Diffusion | Channels / Carrier Proteins | Thermal (Entropy) | Glucose (GLUT), Ions (Channels), Water (Aquaporins) |
| Primary Active | Pumps (ATPases) | ATP Hydrolysis | $Na^+/K^+$ Pump, $Ca^{2+}$ Pump, $H^+$ Pump |
| Secondary Active | Coupled Transport | Ion Gradient | Symporters, Antiporters |
Cotransport Systems (Secondary Active)
These transporters use the potential energy of an ion gradient (usually $Na^+$ in animals, $H^+$ in plants) created by primary pumps to drive the transport of a solute against its gradient.
Image 3: Thermodynamics of Cotransport and Secondary Active Transport

Visual Analysis: Thermodynamics of Cotransport
Symport: SGLT Mechanism (Left)
Detailed look at the Sodium-Glucose Transporter (SGLT):
- Energy Coupling: The $Na^+/K^+$ pump creates a steep $Na^+$ gradient. SGLT uses this gradient energy to "drag" glucose into the cell against its concentration gradient.
- Thermodynamic Profile: Visualized coupling where the favorable free energy change ($\Delta G < 0$) of $Na^+$ movement drives the unfavorable movement ($\Delta G > 0$) of glucose.
Antiport: NHE Mechanism (Right)
Detailed look at the $Na^+$-$H^+$ Exchanger (NHE):
- Molecular Mechanism: Utilizes the $Na^+$ gradient to eject $H^+$ ions from the cell, a critical process for pH regulation.
- Energy Exchange: Similar to symport, the energy of one ion moving down its gradient is harvested to move another ion up its gradient, but in the opposite direction.

Figure 1.2: Symport and Antiport Mechanisms
-
Symport (Cotransport)
Both substances move in the same direction.
Example: SGLT (Sodium-Glucose Transporter) in the intestine/kidney. $Na^+$ moves down its gradient into the cell, dragging Glucose in against its gradient. -
Antiport (Exchanger)
Substances move in opposite directions.
Example: $Na^+$-$H^+$ Exchanger. $Na^+$ enters the cell, while $H^+$ is ejected to regulate intracellular pH.
1.4 Protein Sorting & Signal Hypotheses
Proteins synthesized in the cytosol must be directed to specific organelles. This is achieved via Signal Sequences (amino acid tags).
1. Gated Transport
CYTOSOL ↔ NUCLEUS
Proteins enter through Nuclear Pore Complexes (NPC). They contain a Nuclear Localization Signal (NLS). Key feature: Proteins pass through in a fully folded state.
2. Transmembrane
CYTOSOL → ER / MITO
Proteins utilize protein translocators (e.g., TOM/TIM for mitochondria, Sec61 for ER). Key feature: Proteins must unfold to snake through the channel.
3. Vesicular
ER → GOLGI → PLASMA
Transport via membrane-enclosed vesicles that bud and fuse. The membrane topology (inner vs outer face) is preserved throughout.
The Signal Hypothesis & Co-translational Translocation
Image 4: Co-translational Protein Translocation and Signal Hypothesis

Visual Analysis: Co-translational Translocation
Based on LO 6.3 and 1.4, this visual tracks the journey of a protein from the ribosome to the ER lumen.
- The SRP Cycle: As the ER signal sequence emerges from the ribosome, it is recognized by the SRP (Signal Recognition Particle), which halts translation. The complex then docks onto the SRP Receptor on the Rough ER membrane.
- The Sec61 Translocator: A detailed view of the translocation channel. Key features include the Hydrophobic Plug (sealing the channel when idle), the Lateral Gate (allowing signal sequences to exit into the lipid bilayer), and the central aqueous pore.
- Lumenal Processing: As the polypeptide is threaded through, Signal Peptidase cleaves the signal sequence, and lumenal chaperones guide the protein into its final folded conformation.
Step-by-Step Flow:
- Ribosome begins translation. An ER Signal Sequence (hydrophobic) emerges.
- SRP (Signal Recognition Particle) binds the signal sequence and the ribosome, pausing translation.
- The SRP-Ribosome complex binds to the SRP Receptor on the ER membrane.
- SRP is released. The ribosome passes to a Protein Translocator.
- Translation resumes. The protein is threaded into the ER lumen (Co-translational translocation).
- Signal Peptidase clips off the signal sequence. The protein folds in the ER lumen.
1.5 G-Protein Coupled Receptors (GPCRs)
GPCRs are the largest family of cell-surface receptors (7-pass transmembrane helix). They mediate responses to hormones, neurotransmitters, and light.
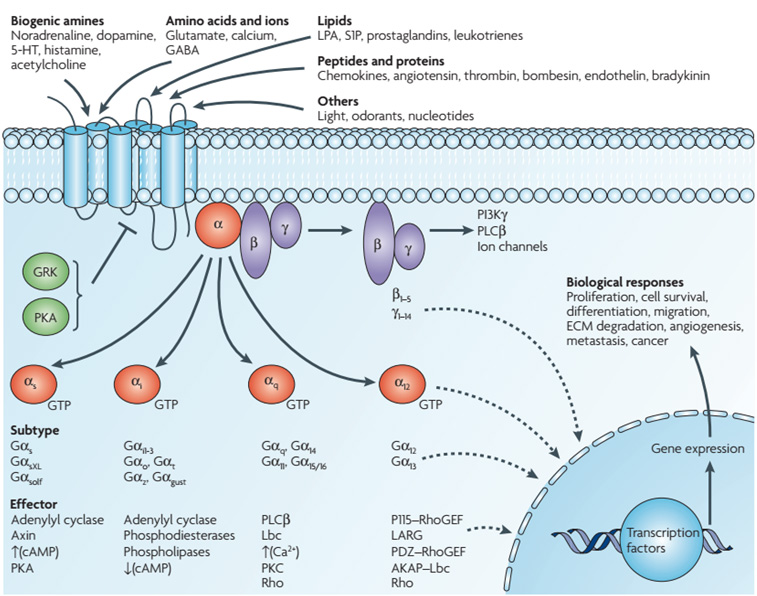
Figure 1.3: Overview of GPCR Activation and Downstream Cascades
Pathway A: Adenylyl Cyclase (Gs)
- Ligand binds GPCR -> Conformational change.
- GPCR acts as GEF for Gs-protein ($\alpha$ subunit exchanges GDP for GTP).
- Activated Gs-$\alpha$ activates Adenylyl Cyclase.
- Enzyme converts ATP into cyclic AMP (cAMP).
- cAMP activates PKA (Protein Kinase A).
- PKA phosphorylates metabolic enzymes (fast) or transcription factors like CREB (slow).
Pathway B: Phospholipase C (Gq)
- Ligand binds GPCR -> Activates Gq-protein.
- Gq activates Phospholipase C (PLC).
- PLC cleaves membrane lipid $PIP_2$ into IP3 (soluble) and DAG (membrane-bound).
- IP3: Binds receptors on Smooth ER -> Releases $Ca^{2+}$ into cytosol.
- DAG + $Ca^{2+}$: Activate PKC (Protein Kinase C).
1.6 Methods & Tools in Biology
Microscopy
- Light Microscope: Resolution ~200nm. Living cells visible. Stains (H&E) needed for contrast.
- Fluorescence Microscope: Uses specific wavelengths to excite fluorophores (e.g., GFP, DAPI). Localization of specific proteins.
- Transmission Electron Microscope (TEM): Resolution ~0.1nm. Electrons pass through specimen. 2D internal ultrastructure.
- Scanning Electron Microscope (SEM): Electrons scatter off surface. 3D topography.
Cell Fractionation
Separation of organelles by centrifugation speed.
- 600 g: Nuclei, Whole cells.
- 15,000 g: Mitochondria, Lysosomes, Peroxisomes.
- 100,000 g: Microsomes (ER/Golgi fragments).
- 300,000 g: Ribosomes, Viruses, Macromolecules.
Epithelial Tissue
Epithelium is a continuous sheet of cells that covers body surfaces, lines cavities, and forms glands. It is avascular, supported by a basement membrane, and exhibits polarity.
Image 5: Classification of Epithelial Tissues and Cell Polarity

Visual Analysis: Epithelial Classification & Polarity
Classification Grid (Left)
A complete visual guide to epithelial types based on layer count and cell shape:
- Layers: Simple (1 layer), Stratified (>1 layer), and Pseudostratified.
- Shapes: Squamous (flat), Cuboidal (cube), Columnar (tall), and Transitional (variable).
- Context: Examples for each type (e.g., Alveoli for Simple Squamous, Trachea for Ciliated Pseudostratified) are integrated.
Cell Polarity & Junctions (Right)
A high-magnification view of a single epithelial cell detailing its functional domains:
- Apical Domain: Features like microvilli (absorption) and cilia (transport).
- Lateral Domain: The "junctional complex" (Tight, Adherens, Desmosomes, Gap) with molecular details like claudins, cadherins, and connexons.
- Basal Domain: Anchoring to the basement membrane (Basal Lamina + Reticular Lamina) via Hemidesmosomes (integrins).
2.1 Classification & Location
| Layers | Shape | Type | Key Locations & Functions |
|---|---|---|---|
| Simple (1 Layer) | Squamous | Simple Squamous | Alveoli, Endothelium, Mesothelium. (Rapid Diffusion/Filtration) |
| Cuboidal | Simple Cuboidal | Kidney Tubules, Thyroid Follicles. (Secretion/Absorption) | |
| Columnar | Simple Columnar | Stomach, Intestine (with Microvilli). (High Absorption/Secretion) | |
| Stratified (>1 Layer) | Squamous | Stratified Squamous | Keratinized: Epidermis (Dry, Waterproof). Non-keratinized: Oral cavity, Vagina (Moist). (Protection) |
| Columnar | Pseudostratified | Respiratory Tract (Trachea). Ciliated. (Mucus Transport) | |
| Variable | Transitional | Bladder, Ureter. Dome cells. (Stretch/Distension) |
2.2 Polarity & Junctions
Intercellular Junctions
- 1. Tight Junction (Zonula Occludens): Most apical. Formed by Claudins/Occludins. Seals paracellular pathway (Barrier function) and maintains polarity.
- 2. Adherens Junction (Zonula Adherens): Below tight junctions. Cadherins link to actin cytoskeleton. Forms an adhesion belt.
- 3. Desmosome (Macula Adherens): Spot weld. Desmogleins link to Intermediate Filaments (Keratin). Resists mechanical stress.
- 4. Gap Junction: Communicating channel formed by Connexons. Allows ions/small molecules to pass. (Heart, Smooth muscle).
Surface Modifications
- Microvilli: Finger-like projections (1-2µm). Core of Actin. Increase surface area for absorption. (Intestine Brush Border).
- Cilia: Hair-like projections (5-10µm). Core of Microtubules (9+2). Motile via dynein. Move mucus. (Trachea, Oviduct).
- Basement Membrane:
1. Basal Lamina (Collagen IV, Laminin - from Epithelium).
2. Reticular Lamina (Collagen III - from Connective).
Anchored by Hemidesmosomes (Integrins).
Connective Tissue
Characterized by sparse cells scattered in an abundant Extracellular Matrix (ECM).
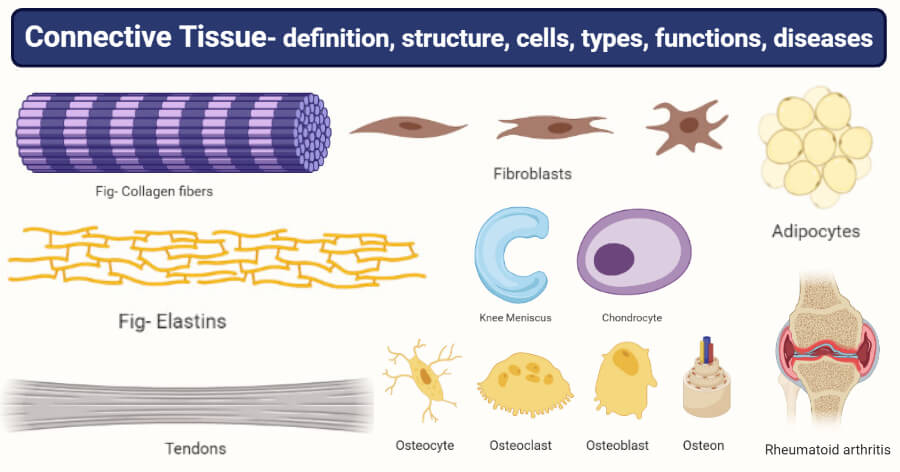
Figure 3.1: Varieties of Connective Tissue
3.1 Extracellular Matrix (ECM)
Protein Fibers
- Collagen Type I: Thick, eosinophilic bundles. High tensile strength. (Bone, Skin, Tendon).
- Collagen Type II: Thin fibrils. (Hyaline Cartilage).
- Collagen Type III (Reticular): Delicate meshwork. (Lymph nodes, Spleen, Liver). Silver stain.
- Elastic Fibers: Elastin core + Fibrillin. Stretch and recoil. (Aorta, Ear).
Ground Substance
A hydrated gel that resists compression.
- GAGs: Long, negative sugar chains (Hyaluronic acid, Chondroitin sulfate). Attract $Na^+$ and water.
- Proteoglycans: Core protein + GAGs (Aggrecan).
- Glycoproteins: Adhesive glue (Fibronectin, Laminin).
3.2 Specialized Tissues
Bone & Cartilage
- Avascular. Cells in Lacunae.
- Hyaline: Type II Collagen. (Joints, Ribs).
- Elastic: Elastic fibers. (Ear).
- Fibrocartilage: Type I. (Discs).
- Calcified Matrix (Hydroxyapatite). Vascular.
- Osteoblasts: Build bone.
- Osteoclasts: Resorb bone (Macrophage origin).
- Osteon: Haversian system in compact bone.
Blood
- Erythrocytes (RBC): 7.5µm. Anucleate biconcave discs. Transport $O_2$. Energy via Anaerobic Glycolysis only.
- Leukocytes (WBC):
Granulocytes: Neutrophil (Bacteria), Eosinophil (Allergy/Parasite), Basophil (Histamine).
Agranulocytes: Lymphocyte (T/B Cells), Monocyte (Macrophage precursor). - Platelets: Thrombocytes. Fragments of Megakaryocytes. Clotting.
Muscle Tissue
Muscle tissue is specialized for contraction via the interaction of Actin and Myosin filaments. It consumes ATP to generate force.

Figure 4.1: Muscle Hierarchy (Muscle -> Fascicle -> Fiber -> Myofibril -> Sarcomere)

Figure 4.2: Fiber Anatomy (Sarcolemma, T-tubules, SR)
4.1 Classification
| Type | Striations | Control | Structure | Features |
|---|---|---|---|---|
| Skeletal | Yes | Voluntary | Cylindrical, Multinucleated (peripheral) | Triads (T-tubule + 2 SR). Troponin C. |
| Cardiac | Yes | Involuntary | Branched, 1-2 Nuclei (central) | Intercalated Discs (Gap junctions + Desmosomes). Diads. |
| Smooth | No | Involuntary | Fusiform, 1 Nucleus (central) | Dense Bodies (no sarcomeres). Calmodulin (no Troponin). |
4.2 The Sarcomere & Contraction
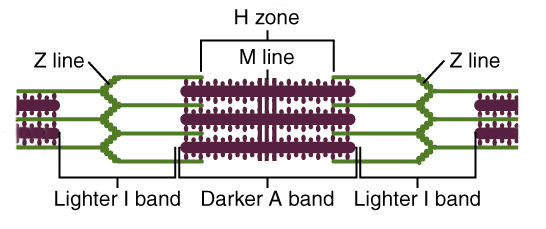
Figure 4.3: The Functional Unit of Contraction (Z-line to Z-line)
Band Changes
- A Band: Length of myosin. Remains constant.
- I Band: Actin only. Shortens.
- H Zone: Myosin only. Shortens.
- Z Lines: Move closer together.
Excitation-Contraction Coupling
- Nerve AP releases ACh -> Muscle AP.
- AP travels down T-tubules to Triad.
- Voltage sensor (DHP) opens Ryanodine Receptors (RyR) on SR.
- $Ca^{2+}$ floods sarcoplasm.
- $Ca^{2+}$ binds Troponin C.
- Troponin moves Tropomyosin off actin sites.
- Myosin heads bind actin (Crossbridge) -> Power Stroke (ATP needed for release).
Nervous Tissue
Nervous tissue allows for rapid communication and integration. It consists of excitable Neurons and supporting Neuroglia.

Figure 5.1: Functional Anatomy of a Neuron
5.1 Physiology: The Action Potential
The fundamental unit of neural communication.

Figure 5.2: Phases of Membrane Potential
- Resting (-70mV): Established by $Na^+/K^+$ pump and $K^+$ leak channels. Inside is negative.
- Depolarization: Threshold reached. Voltage-gated Na+ Channels open. $Na^+$ rushes IN. Potential shoots to +30mV.
- Repolarization: $Na^+$ channels inactivate. Voltage-gated K+ Channels open. $K^+$ rushes OUT. Potential drops.
- Hyperpolarization: $K^+$ channels constitute slow closing. Potential dips below resting level.
- Refractory Period: Ensures one-way propagation.
5.2 Synaptic Transmission
- Action potential arrives at the Axon Terminal.
- Depolarization opens Voltage-gated $Ca^{2+}$ Channels.
- $Ca^{2+}$ influx triggers synaptic vesicles (SNARE proteins) to fuse with membrane.
- Neurotransmitter (NT) is released into the cleft (Exocytosis).
- NT binds to Ligand-gated ion channels on postsynaptic cell.
- Channels open -> Ion flux -> EPSP or IPSP.

Figure 5.3: The Chemical Synapse
5.3 Neuroglia (Support Cells)
CNS Glia
- Astrocytes: Blood-Brain Barrier, Metabolic support, K+ buffering.
- Oligodendrocytes: Myelinate multiple axons.
- Microglia: Immune defense (Phagocytes).
- Ependymal Cells: Line ventricles, produce CSF.
PNS Glia
- Schwann Cells: Myelinate a single axon segment. Aid regeneration.
- Satellite Cells: Support neuron cell bodies in ganglia.