The Molecular Logic
of Living Organisms
A definitive, uncompromising exploration of Lipids, Carbohydrates, Amino Acids, Proteins, Enzymes, DNA/RNA Structure, and Molecular Genetics. Based strictly on the fundamental principles of Lehninger Biochemistry.
The Molecular Logic of Life
Biochemistry seeks to describe in molecular terms the structures, mechanisms, and chemical processes shared by all organisms. It provides the ultimate physical explanation for the remarkable properties of living systems.
The Solvent of Life: Water
Water is the essential matrix of life. Its polar nature and ability to form hydrogen bonds dictate the folding of proteins, the assembly of membranes, and the solvation of biomolecules.
Visualizing Solvation (Hydration Process)

Water molecules surround ions based on charge. The oxygen atoms ($\delta-$) orient towards cations, while the hydrogen atoms ($\delta+$) face anions, creating a hydration shell that stabilizes solutes.
Thermodynamics & Transport
Life requires a constant input of energy and the efficient transport of matter. Organisms must overcome physical barriers to maintain homeostasis across microscopic and macroscopic scales.
Logistics of Life: Diffusion vs. Bulk Transport

Simple diffusion ($t \propto x^2$) is only effective over microscopic distances. Multicellular life requires bulk transport (circulatory systems) to deliver nutrients and oxygen to deep tissues efficiently.
Course Architecture
Comprehensive Outline of the Lehninger Biochemistry Masterclass
Module 1: Lipids
- 1.1 Lipid Classification & The Lipidome
- 1.2 Saturated vs Unsaturated Fatty Acids
- 1.3 Cis/Trans Geometry & Melting Points
- 1.4 Triacylglycerols & Energy Storage
- 1.5 Glycerophospholipids & Sphingolipids
- 1.6 Sterols, Cholesterol & Fluidity Buffering
- 1.7 Amphipathic Aggregation (Micelles/Liposomes)
- 1.8 Eicosanoids (Prostaglandins, Thromboxanes)
- 1.9 Fat-Soluble Vitamins (A, D, E, K)
Module 2: Carbohydrates
- 2.1 Aldoses vs Ketoses (Trioses to Hexoses)
- 2.2 Stereochemistry, Enantiomers & Epimers
- 2.3 Cyclic Structures (Hemiacetals/Hemiketals)
- 2.4 Haworth Perspectives & α/β Anomers
- 2.5 Mutarotation & Reducing Sugars
- 2.6 Disaccharides & O-Glycosidic Linkages
- 2.7 Starch (Amylose/Amylopectin) & Glycogen
- 2.8 Structural Polysaccharides (Cellulose, Chitin)
- 2.9 Glycosaminoglycans (GAGs) & The ECM
Module 3: Amino Acids
- 3.1 The 20 Standard Amino Acids
- 3.2 Nonpolar & Aliphatic R Groups
- 3.3 Aromatic R Groups & UV Absorbance (A280)
- 3.4 Polar Uncharged Groups & Disulfide Bridges
- 3.5 Acidic & Basic Charged R Groups
- 3.6 Buffers & Henderson-Hasselbalch Equation
- 3.7 Isoelectric Point (pI) & Electrophoresis
- 3.8 Peptide Bond Condensation Reaction
- 3.9 Evolutionary Bioinformatics & Homologs
Module 4: Protein 3D Structure
- 4.1 Hierarchical Levels (1°, 2°, 3°, 4°)
- 4.2 Rigidity & Resonance of the Peptide Bond
- 4.3 Dihedral Angles (φ, ψ) & Ramachandran Plot
- 4.4 Secondary Structures (α-Helix, β-Sheet)
- 4.5 Fibrous Proteins (α-Keratin, Collagen, Silk)
- 4.6 Globular Proteins, Motifs & Domains
- 4.7 Thermodynamics & Levinthal's Paradox
- 4.8 Molecular Chaperones (Hsp70, GroEL/GroES)
- 4.9 Pathological Amyloidosis & Prion Diseases
Module 5: Protein Function
- 5.1 Reversible Ligand Binding & Induced Fit
- 5.2 Oxygen Binding & The Heme Prosthetic Group
- 5.3 Distal Histidine & Carbon Monoxide Shielding
- 5.4 Myoglobin vs Hemoglobin Architecture
- 5.5 T-State vs R-State Conformational Shifts
- 5.6 The Hill Equation & Positive Cooperativity
- 5.7 Allosteric Regulation: The Bohr Effect
- 5.8 2,3-Bisphosphoglycerate (BPG) Modulation
- 5.9 Sickle Cell Anemia: Molecular Pathology
Module 6: Enzymes
- 6.1 Biological Catalysts & Activation Energy (ΔG‡)
- 6.2 Reaction Coordinate Diagrams
- 6.3 Induced Fit vs. Lock and Key Models
- 6.4 Cofactors, Coenzymes & Specific Activity
- 6.5 Michaelis-Menten Kinetics (Vmax, Km, kcat)
- 6.6 Competitive & Uncompetitive Inhibition
- 6.7 Mixed (Non-competitive) Inhibition
- 6.8 Allosteric Regulation & Covalent Modification
- 6.9 Zymogens & Proteolytic Cleavage
Module 7: Nucleic Acids
- 7.1 Components: Phosphate, Pentose & Bases
- 7.2 Nucleosides vs Nucleotides
- 7.3 Purines (A, G) vs Pyrimidines (C, T, U)
- 7.4 Phosphodiester Linkages & Directionality
- 7.5 The Watson-Crick DNA Double Helix
- 7.6 Antiparallel Strands & Base Pairing Rules
- 7.7 Base Stacking & Thermodynamic Stability
- 7.8 Major/Minor Grooves & Recognition
- 7.9 Reversible Denaturation & Melting Temp (Tm)
Module 8: Genes & Chromosomes
- 8.1 Gene Definitions & Biological Products
- 8.2 Eukaryotic Architecture: Introns vs Exons
- 8.3 DNA Topology & Linking Number (Lk)
- 8.4 Supercoiling: Twist (Tw) & Writhe (Wr)
- 8.5 Topoisomerases & Structural Regulation
- 8.6 Eukaryotic Chromatin Organization
- 8.7 The Nucleosome & Histone Octamer Core
- 8.8 Electrostatic Neutralization by Arg/Lys
- 8.9 Linker DNA (H1) & Higher-Order Compaction
Module 9: DNA Metabolism
- 9.1 The Thermodynamic Cost of Absolute Fidelity
- 9.2 Semiconservative Replication Dynamics
- 9.3 Bidirectional Synthesis & Replication Forks
- 9.4 Semi-discontinuous Synthesis
- 9.5 Leading vs Lagging Strands (Okazaki Fragments)
- 9.6 The Replisome: DNA Helicase & SSB Proteins
- 9.7 DNA Primase & Initial RNA Primers
- 9.8 DNA Polymerase III (Synthesis & Proofreading)
- 9.9 Primer Removal (DNA Pol I) & DNA Ligase
Module 10: RNA Metabolism
- 10.1 Transcription & DNA-Dependent RNA Polymerase
- 10.2 Promoters & Transcription Initiation
- 10.3 Template Strand vs Coding (Non-template) Strand
- 10.4 Eukaryotic Pre-mRNA Maturation Process
- 10.5 5' Capping (7-methylguanosine)
- 10.6 The Spliceosome & Intron Removal
- 10.7 3' Polyadenylation (Poly-A Tail)
Module 11: Protein Metabolism
- 11.1 The Universal Genetic Code & Codons
- 11.2 Code Properties (Unambiguous, Degenerate)
- 11.3 The Wobble Hypothesis
- 11.4 Activation of Amino Acids
- 11.5 Aminoacyl-tRNA Synthetases (The 2nd Code)
- 11.6 Ribosome Structure (Large & Small Subunits)
- 11.7 Translation Initiation & fMet/Met
- 11.8 Elongation, Translocation & Peptide Bonding
- 11.9 Termination via Release Factors
Module 12: Neurogenesis
- 12.1 Adult Hippocampal Neurogenesis Paradigm
- 12.2 The Dentate Gyrus & Subgranular Zone (SGZ)
- 12.3 Neural Stem Cells & Cellular Proliferation
- 12.4 Behavioral Regulation (Exercise vs Stress)
- 12.5 The Endocannabinoid System (ECS)
- 12.6 CB1/CB2 Receptors & Retrograde Signaling
- 12.7 Pharmacological Modulation & Plasticity
- 12.8 Anxiolytic & Antidepressant Effects
Organic Chemistry
Structure, Stereochemistry, Mechanisms & Synthesis
Module 13: Bonding & Hybridization
Module 14: Isomers & Forces
Module 16: Conformations
Module 17: Cycloalkanes
Module 19: Alkenes
Module 20: Alcohols & Amines
Module 22: Substitution
Module 23: Elimination
Module 25: Aromaticity
Module 26: Carbonyls
Module 1: Lipids
Energy Storage, Membrane Architecture, and Signaling
Lipids constitute a highly heterogeneous, structurally diverse group of cellular molecules fundamentally characterized by their extreme insolubility in water and high solubility in nonpolar organic solvents (like chloroform or ether). Unlike proteins, nucleic acids, and polysaccharides, lipids are strictly not polymeric. Instead, they aggregate noncovalently driven primarily by the hydrophobic effect to form enormous macromolecular complexes, such as biological lipid bilayers and immense intracellular lipid droplets. The complete, specific complement of lipids present in a highly specific cell type under particular physiological conditions is systematically defined as the lipidome.
Physiological Logistics: Lipid Digestion and Absorption

Because lipids are insoluble in the aqueous environment of the digestive tract, their digestion requires specialized processing. This diagram illustrates the role of bile salts in emulsification (increasing surface area), the formation of mixed micelles for transport to the intestinal mucosa, and the intracellular assembly of chylomicrons for transport into the lymphatic system via lacteals.
| Functional Category | Lipid Class | Core Backbone Component | Biological Significance & Function |
|---|---|---|---|
| Storage Lipids | Triacylglycerols (Fats) | Glycerol | Primary long-term energy depot. Highly reduced carbon yields maximal ATP upon complete oxidation. Stored completely unhydrated in adipocytes. |
| Waxes | Long-chain Alcohol | Esters of long-chain fatty acids with long-chain alcohols. Provide extreme water repellency for feathers, leaves, and structural rigidity (honeycombs). | |
| Structural Membrane Lipids | Glycerophospholipids | Glycerol 3-Phosphate | Absolute foundation of all eukaryotic cell membranes. Amphipathic geometry forces spontaneous lipid bilayer assembly in aqueous environments. |
| Sphingolipids | Sphingosine | Crucial in neural tissues (myelin sheath). Carbohydrate-bearing sphingolipids (glycosphingolipids) strictly determine ABO human blood types. | |
| Sterols (e.g., Cholesterol) | Steroid Nucleus (4 fused rings) | Rigid, planar structural buffers. Modulate membrane fluidity across various temperatures. Direct precursors for all steroid hormones and bile acids. | |
| Signaling Lipids | Eicosanoids (PGs, TXs, LTs) | Arachidonic Acid (20:4) | Potent paracrine hormones. Mediate aggressive inflammatory responses, pain induction, fever generation, and critical blood clotting cascades. |
1.1 Fatty Acids: Structure, Nomenclature, and Physical Properties
Library of Common Medical & Biological Lipids

A comprehensive reference for the structures and common names of medically significant organic compounds, including fatty acids and their functional derivatives.
Fatty acids are the fundamental building blocks of most saponifiable lipids. They are essentially water-insoluble aliphatic hydrocarbons terminating in a highly polar carboxylic acid group ($-COOH$). Because they are synthesized in biological systems primarily by the successive condensation of two-carbon acetate units, the vast majority of naturally occurring fatty acids possess an unbranched chain with an even number of carbon atoms, typically ranging exclusively from 12 to 24.
Saturated Fatty Acids
Saturated fatty acids contain absolutely no carbon-carbon double bonds in their extended hydrocarbon tail. They are completely "saturated" with hydrogen atoms.
- Conformation: Because rotation is completely free around every single carbon-carbon bond, the molecule thermodynamically prefers the fully extended, linear zigzag conformation to minimize steric repulsion.
- Packing & Melting Point: These linear, unbranched chains can pack incredibly tightly together into highly ordered, quasi-crystalline arrays. This tight packing maximizes stabilizing intermolecular van der Waals interactions.
- Result: Consequently, saturated fatty acids require massive thermal energy to disrupt their crystalline lattice, giving them extraordinarily high melting points. They are almost universally solid or waxy at human room temperature (e.g., animal fats, butter, tallow).
- Palmitic Acid (16:0)
- Stearic Acid (18:0)
- Arachidic Acid (20:0)
Unsaturated Fatty Acids
Unsaturated fatty acids contain one (monounsaturated) or multiple (polyunsaturated) carbon-carbon double bonds.
- Cis Configuration: In nearly all naturally occurring unsaturated fatty acids, the double bonds are strictly in the cis geometric configuration.
- The Structural Kink: A cis double bond introduces a rigid, unyielding ~30-degree "kink" or bend directly into the aliphatic hydrocarbon chain.
- Packing & Melting Point: This permanent structural bend completely prevents the chains from aligning linearly and packing tightly together. The resulting poor packing drastically weakens intermolecular van der Waals interactions.
- Result: Consequently, they require vastly less thermal energy to separate, resulting in remarkably low melting points. They are characteristically liquids (oils) at room temperature (e.g., olive oil, sunflower oil).
- Oleic Acid (18:1 $\Delta^9$)
- Linoleic Acid (18:2 $\Delta^{9,12}$)
- Arachidonic Acid (20:4 $\Delta^{5,8,11,14}$)
Atomic Architecture: Saturated, Cis-Unsaturated, and Trans-Unsaturated Fatty Acids
Commercially, liquid vegetable oils (rich in cis-unsaturated fats) are subjected to extreme industrial partial hydrogenation to convert them into solid, spreadable fats (like margarine) and drastically increase their shelf life by preventing oxidative rancidity. This violent chemical process converts many cis double bonds into single bonds. However, it inadvertently causes a side reaction that isomerizes remaining cis double bonds into trans double bonds.
As structurally illustrated above, a trans double bond completely lacks the rigid 30° kink of a cis bond. Instead, it allows the fatty acid to assume an extended, linear conformation that is physically virtually identical to a saturated fatty acid. Consequently, trans fats can pack tightly, giving them higher melting points. Biologically, dietary intake of trans fats is disastrous; they powerfully raise LDL (low-density lipoprotein, "bad" cholesterol) levels and lower HDL (high-density lipoprotein, "good" cholesterol) levels, promoting massive systemic inflammatory responses and drastically elevating the risk of coronary heart disease.
You will frequently be asked to order fatty acids by melting point on biochemistry exams. Apply this exact logical algorithm:
- Count the double bonds (Degree of Unsaturation): This is the absolute dominant factor. The more cis double bonds present, the lower the melting point. A polyunsaturated fat (e.g., Arachidonic acid, 20:4) will have a massively lower melting point than a monounsaturated fat (e.g., Oleic acid, 18:1), which is lower than any saturated fat.
- Assess the Geometry (Cis vs Trans): If comparing an 18:1 cis acid (Oleic acid) against an 18:1 trans acid (Elaidic acid), the trans isomer mimics a saturated fat, packing tighter, resulting in a significantly higher melting point.
- Analyze the Chain Length (Only if unsaturation is equal): If comparing two fully saturated fats, the longer the hydrocarbon chain, the greater the total surface area for stabilizing van der Waals interactions, and therefore, the higher the melting point (e.g., Arachidic acid 20:0 > Stearic acid 18:0 > Palmitic acid 16:0).
Comprehensive Lipid Nomenclature & Properties
| Carbon Skeleton | Common Name | Systematic Name | Melting Point (°C) | Biological Source / Significance |
|---|---|---|---|---|
| 12:0 | Lauric acid | Dodecanoic acid | 44.2 | Coconut oil, palm kernel oil. |
| 14:0 | Myristic acid | Tetradecanoic acid | 53.9 | Nutmeg, dairy products. |
| 16:0 | Palmitic acid | Hexadecanoic acid | 63.1 | Palm oil, animal fats; most common saturated FA. |
| 18:0 | Stearic acid | Octadecanoic acid | 69.6 | Animal fats, cocoa butter. |
| 20:0 | Arachidic acid | Eicosanoic acid | 76.5 | Peanut oil. |
| 16:1($\Delta^{9}$) | Palmitoleic acid | 9-Hexadecenoic acid | -0.5 | Macadamia oil, ubiquitous in animal tissues. |
| 18:1($\Delta^{9}$) | Oleic acid | 9-Octadecenoic acid | 13.4 | Olive oil; most common unsaturated FA. |
| 18:2($\Delta^{9,12}$) | Linoleic acid | 9,12-Octadecadienoic acid | -5.0 | Omega-6 essential fatty acid; corn, soy oils. |
| 18:3($\Delta^{9,12,15}$) | $\alpha$-Linolenic acid | 9,12,15-Octadecatrienoic acid | -11.0 | Omega-3 essential fatty acid; flaxseed, chia. |
| 20:4($\Delta^{5,8,11,14}$) | Arachidonic acid | 5,8,11,14-Eicosatetraenoic acid | -49.5 | Precursor to eicosanoid hormones. |
1.2 Triacylglycerols (Neutral Fats) and Energy Storage
Triacylglycerols (also known as triglycerides) are the simplest lipids constructed from fatty acids. They consist of exactly three fatty acid molecules covalently linked to a single, central glycerol backbone via ester bonds. Because the polar hydroxyls of glycerol and the polar carboxylates of the fatty acids are entirely consumed in ester bond formation, triacylglycerols are completely nonpolar, highly hydrophobic, and fundamentally insoluble in water. They are scientifically classified as "neutral fats" because they bear absolutely no ionic charge at biological pH.
The Thermodynamics of Fat Storage
Vertebrates store immense quantities of triacylglycerols in specialized cells called adipocytes (fat cells), where massive lipid droplets occupy nearly the entire cellular volume. Plant seeds also store huge amounts of oils to provide energy during germination. Triacylglycerols possess two massive evolutionary advantages over polysaccharides (like glycogen or starch) as long-term energy storage depots:
- Superior Energy Yield: The carbon atoms in aliphatic fatty acid chains are highly reduced (mostly $-CH_2-$). Complete enzymatic oxidation of these chains yields more than twice the metabolic energy per gram (~9 kcal/g) compared to the more oxidized carbons of carbohydrates or proteins (~4 kcal/g).
- Zero Hydration Weight: Because triacylglycerols are immensely hydrophobic, they are stored completely unhydrated. In stark contrast, polysaccharides are highly polar and rigidly bind roughly 2 grams of water per gram of carbohydrate. An organism storing energy solely as glycogen would be impossibly heavy and unable to move efficiently.
Condensation: Triacylglycerol Synthesis
Structural Backbones: Glycerol vs. Sphingosine
1.3 Structural Membrane Lipids: The Amphipathic Imperative
While storage lipids are entirely nonpolar, the structural lipids that construct cellular membranes are definitively amphipathic. This means they possess a dual chemical nature: one end of the molecule consists of a massively bulky, hydrophobic tail region (usually two long fatty acid chains), while the opposite end consists of a highly polar or electrically charged hydrophilic head group. In the aqueous environment of a living cell, these physical properties mathematically force these lipids to spontaneously self-assemble into a highly stable lipid bilayer, the absolute fundamental architectural basis of all biological membranes.
The Fluid Mosaic Model proposed by Singer and Nicolson dictates that this bilayer is not rigid but acts as a dynamic two-dimensional fluid. However, specialized microdomains called Lipid Rafts—heavily enriched in tightly packed sphingolipids and cholesterol—float within this fluid, serving as highly organized organizing centers for massive signal transduction cascades.
1. Glycerophospholipids
The paramount, most abundant lipids in eukaryotic cell membranes. They are strictly built upon a core scaffold of L-glycerol 3-phosphate.
- C-1: Typically esterified to a saturated fatty acid (e.g., Palmitate).
- C-2: Typically esterified to a cis-unsaturated fatty acid (e.g., Oleate, creating a necessary kink for fluidity).
- C-3: Attached via a phosphodiester linkage to a highly polar head group.
- Classes: The specific head group entirely dictates the class. Choline yields Phosphatidylcholine (Lecithin). Ethanolamine yields Phosphatidylethanolamine. Serine yields Phosphatidylserine (which carries a net negative charge and signals apoptosis when exposed on the outer leaflet).
2. Sphingolipids & Sterols
Massively important membrane components structurally distinct from glycerophospholipids.
- Sphingosine Base: Sphingolipids are built on a long-chain amino alcohol. Their core unit is Ceramide (fatty acid linked via an amide bond).
- Glycosphingolipids: Contain complex sugar head groups linked via glycosidic bonds (no phosphate). They strictly determine human blood groups (A, B, O) on the erythrocyte surface.
- Cholesterol: The rigid core consists of four fused planar rings. It functions as a master Fluidity Buffer: At high temps, its rigid rings restrict fatty acid motion (decreasing fluidity). At low temps, its bulky structure prevents fatty acid crystallization (maintaining fluidity).
Thermodynamics of Amphipathic Lipid Aggregation in Water
A common exam question presents various lipid structures and asks which possess "detergent activity" (the ability to emulsify oils into water).
- To act as a detergent, a molecule must be strictly amphipathic with a single hydrophobic tail (wedge-shaped geometry), which forces the spontaneous formation of micelles in water.
- Example 1: Potassium hexadecanoate ($CH_3(CH_2)_{14}COO^-K^+$). This is a classic soap (a fatty acid salt). It has a highly charged ionic head (carboxylate) and exactly one long hydrophobic tail. It forms micelles. YES, it has detergent activity.
- Example 2: 1-Hexadecanol ($CH_3(CH_2)_{15}OH$). While it has a polar hydroxyl group, it is uncharged and entirely too weakly polar to overcome the massive hydrophobic effect of the long 16-carbon chain to form stable micelles in water. NO, it is insoluble and lacks detergent activity.
- Example 3: A standard phospholipid. Phospholipids have a highly charged head group but possess TWO massive fatty acid tails (cylindrical geometry). Because they are cylindrical, they are physically incapable of forming tight, spherical micelles. Instead, they form extended bilayers. NO, standard phospholipids are poor detergents.
1.4 Active Lipids: Signals, Cofactors, and Pigments
Principle : While storage lipids (triacylglycerols) and structural membrane lipids constitute the overwhelming bulk of cellular lipid mass, a remarkable variety of lipids present in minute quantities play absolutely indispensable roles as active biochemical signals, crucial enzyme cofactors, and light-absorbing biological pigments.
Eicosanoids (Paracrine Signals)
Eicosanoids are incredibly potent paracrine hormones derived directly from the enzymatic oxidation of arachidonic acid (a 20-carbon polyunsaturated fatty acid). They act strictly locally, on cells near the point of their synthesis.
- Prostaglandins (PGs): Contain a five-membered ring. They intensely regulate inflammation, induce fever, modulate pain perception, and stimulate smooth muscle contraction (e.g., during menstruation and labor).
- Thromboxanes (TXs): Contain a six-membered cyclic ether ring. Produced primarily by platelets, they act as powerful vasoconstrictors and facilitate the formation of blood clots. Nonsteroidal anti-inflammatory drugs (NSAIDs) like aspirin heavily inhibit the cyclooxygenase enzyme required to synthesize PGs and TXs.
- Leukotrienes (LTs): Characterized by three conjugated double bonds. They are profound biological signals; overproduction dramatically induces smooth muscle contraction in the airways, precipitating severe asthmatic attacks.
Steroid Hormones
Steroid hormones are heavily oxidized, functional derivatives of sterols. They inherently lack the extended alkyl chain attached to ring D of cholesterol, rendering them significantly more polar than cholesterol itself.
- They are transported systematically through the bloodstream securely bound to specific carrier proteins.
- Upon reaching target tissues, they pass freely through the plasma membrane to bind highly specific receptor proteins located in the nucleus.
- The hormone-receptor complex acts as a powerful transcription factor, triggering profound changes in gene expression and global metabolism.
- Major examples include male and female sex hormones (Testosterone, Estradiol) and hormones produced by the adrenal cortex (Cortisol regulating glucose metabolism, Aldosterone regulating salt excretion).
Fat-Soluble Vitamins (A, D, E, K)
Vitamins are essential organic compounds that cannot be synthesized by vertebrates and must be obtained via the diet. The fat-soluble vitamins are all structurally classified as isoprenoid compounds.
- Vitamin A (Retinol): The crucial precursor for the visual pigment retinal, essential for phototransduction in the eye. Also vital for epithelial tissue maintenance.
- Vitamin D (Cholecalciferol): Formed in the skin via UV irradiation of 7-dehydrocholesterol. It is metabolically converted into a hormone that precisely regulates calcium and phosphorus uptake in the intestine and deposition in bone.
- Vitamin E (Tocopherols): Possess a substituted aromatic ring and a long isoprenoid side chain. They function as profound biological antioxidants, actively scavenging highly reactive free radicals to prevent devastating oxidative damage to membrane lipids.
- Vitamin K: Absolutely essential for the post-translational activation of prothrombin and other crucial blood-clotting factors.
Because lipids are insoluble in water, their extraction and fractionation require the use of organic solvents and specialized chromatographic techniques.
- Lipid Extraction: Tissues are typically homogenized in a mixture of chloroform, methanol, and water. This creates a two-phase system: proteins and sugars partition into the upper aqueous methanol phase, while all neutral and polar lipids partition strictly into the lower dense chloroform phase.
- Adsorption Chromatography: The lipid mixture is applied to a column of insoluble polar material (like silica gel). Neutral lipids (triacylglycerols, sterols) do not bind and elute first with nonpolar solvents (chloroform). Polar lipids (phospholipids) bind tightly to the silica and are eluted subsequently using more polar solvents (methanol).
- Thin-Layer Chromatography (TLC): Utilizing a glass plate coated with silica gel, lipid mixtures are separated based on their relative polarities. Less polar lipids travel furthest up the plate with the advancing solvent front.
- Mass Spectrometry (MS): Allows the precise, definitive structural identification of individual lipid molecules, revealing exact chain length, degree of saturation, and exact placement of double bonds without prior degradation.
Module 2: Carbohydrates & Glycobiology
Energy Metabolism, Structural Matrices, and Cellular Recognition
Carbohydrates (sugars) are definitively the most abundant biomolecules on planet Earth. Structurally, they are formally defined as polyhydroxy aldehydes or polyhydroxy ketones (or substances that completely yield such compounds upon acid hydrolysis). They serve as immediate energy sources, rigid structural elements in plant cell walls and animal connective tissues, and highly informative molecular tags playing central, irreplaceable roles in complex signaling and precise cell-cell recognition events. The comprehensive, systematic study of all complex carbohydrate components of a given cell or tissue is strictly defined as Glycomics.
2.1 Monosaccharides and Stereochemistry
Principle 1: Monosaccharides are the simplest sugars, consisting of a single polyhydroxy aldehyde or ketone unit. Because they inherently contain one or more chiral carbon centers, they exist as multiple distinct stereoisomers, requiring precise nomenclature.
| Classification | Common Name | Chemical Formula | Biological Context & Structural Significance |
|---|---|---|---|
| Aldotriose | D-Glyceraldehyde | $C_3H_6O_3$ | The simplest aldose and the universal reference point for D/L-isomerism. It is a critical intermediate in the glycolytic pathway as Glyceraldehyde-3-phosphate (G3P). |
| Ketotriose | Dihydroxyacetone | $C_3H_6O_3$ | The simplest ketose. It is the only monosaccharide without a chiral center. Its phosphorylated form (DHAP) is in rapid equilibrium with G3P during glycolysis via the enzyme triose phosphate isomerase. |
| Aldopentose | D-Ribose | $C_5H_{10}O_5$ | The structural backbone of RNA, ATP, and NAD+/NADP+. In its deoxygenated form (2-Deoxy-D-ribose), it forms the fundamental stable framework of DNA. |
| Aldohexose | D-Glucose Dextrose / Blood Sugar |
$C_6H_{12}O_6$ | The absolute primary fuel source for the human brain and erythrocytes. It exists predominantly in the pyranose form. It is the fundamental monomer of starch, cellulose, and glycogen. |
| Ketohexose | D-Fructose Levulose / Fruit Sugar |
$C_6H_{12}O_6$ | The sweetest naturally occurring sugar. It bypasses the key regulatory step of glycolysis (PFK-1) when metabolized in the liver, which contributes to rapid lipogenesis. |
| Aldohexose | D-Galactose | $C_6H_{12}O_6$ | A C-4 epimer of glucose. It is essential for the synthesis of lactose in mammary glands and is a key component of glycolipids and glycoproteins in neural cell membranes. |
Aldoses vs. Ketoses
Based on the specific location of the carbonyl group, simple sugars fall into two major chemical families:
|
H-C-OH
|
R
|
C=O
|
R
- Epimers: Two sugars that differ solely in the absolute configuration around exactly one specific chiral carbon atom. For example, D-Glucose and D-Galactose are C-4 epimers; D-Glucose and D-Mannose are C-2 epimers.
- Enantiomers: Non-superimposable mirror-image stereoisomers (D vs L forms). By convention, the D/L designation is strictly determined by the configuration of the chiral center furthest from the carbonyl carbon, using D-glyceraldehyde as the reference standard. The vast majority of naturally occurring hexoses are D-isomers.
Cyclic Structures & Anomers
In aqueous solution, aldotetroses and all larger monosaccharides occur primarily not as open chains, but as stable cyclic ring structures. The carbonyl group undergoes an intramolecular covalent reaction with the oxygen of a hydroxyl group along the chain, forming a stable hemiacetal (from an aldose) or hemiketal (from a ketose).
- Pyranoses and Furanoses: Six-membered ring compounds are termed pyranoses (resembling pyran), whereas five-membered rings are termed furanoses (resembling furan). D-Glucose preferentially forms a glucopyranose ring.
- The Anomeric Carbon: The former carbonyl carbon intimately involved in hemiacetal formation becomes a new chiral center, designated the anomeric carbon.
- $\alpha$ and $\beta$ Anomers: Isomeric forms differing solely in the configuration at the anomeric carbon. In a Haworth perspective drawing, if the anomeric hydroxyl ($-\text{OH}$) group points in the opposite direction (trans) relative to the $-\text{CH}_2\text{OH}$ reference group (C-6), it is designated the $\alpha$ anomer. If it points in the same direction (cis), it is the $\beta$ anomer.
- Mutarotation: The spontaneous interconversion of $\alpha$ and $\beta$ anomers in aqueous solution via the transient open-chain intermediate form. A solution of pure $\alpha$-D-glucose will mutate over time to reach an equilibrium mixture containing roughly one-third $\alpha$-D-glucose and two-thirds $\beta$-D-glucose.
Haworth Perspectives: $\alpha$ vs $\beta$ Anomers of D-Glucopyranose
Fischer Projections: Hexose Stereochemistry & Epimers
2.2 Cyclic Structures: Hemiacetals, Hemiketals, and Anomers
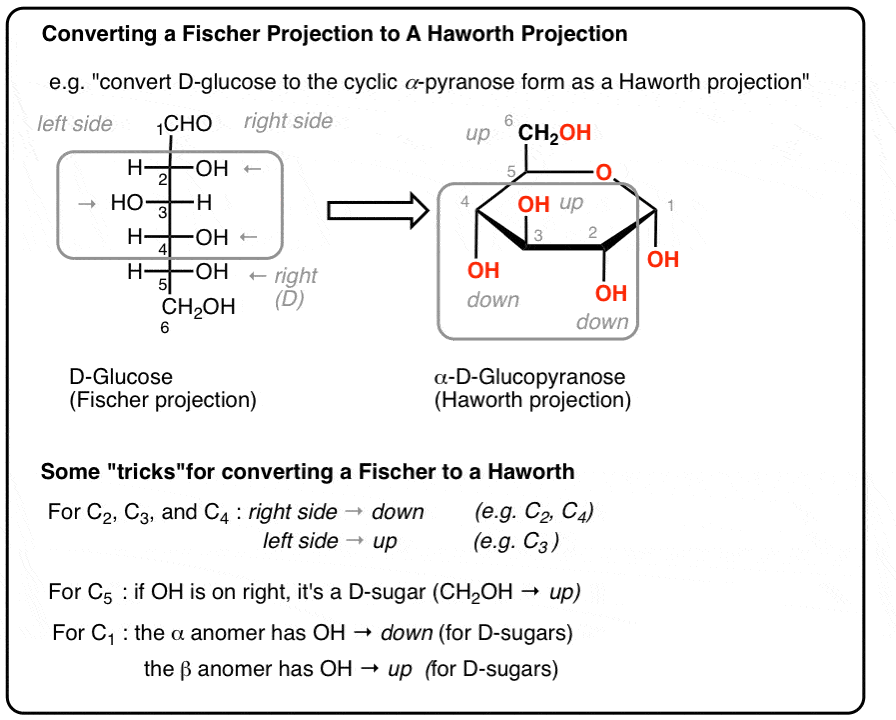
Animated conversion from Fischer projection to Haworth projection

Fischer vs Haworth projection conversion diagram for monosaccharides
In aqueous biological solutions, aldotetroses and all larger monosaccharides simply do not exist as straight open chains. The highly electrophilic carbonyl group undergoes an explosive intramolecular nucleophilic attack by the oxygen of a distal hydroxyl group located along the same chain. This creates a highly stable cyclic ring structure: a hemiacetal (derived from an aldose) or a hemiketal (derived from a ketose).
Anomeric Carbons and $\alpha$/$\beta$ Designation
The original former carbonyl carbon that was intimately involved in hemiacetal formation suddenly becomes a completely new, extra chiral center. This specific carbon is permanently designated the anomeric carbon.
- $\alpha$ Anomer: In a standard Haworth perspective drawing, if the newly formed anomeric hydroxyl ($-OH$) group points in the completely opposite direction (trans) relative to the $-CH_2OH$ reference group (usually C-6), it is specifically designated the $\alpha$ anomer.
- $\beta$ Anomer: If the anomeric hydroxyl points in the exact same direction (cis) as the $-CH_2OH$ reference group, it is strictly designated the $\beta$ anomer.
- Mutarotation: In solution, the $\alpha$ and $\beta$ anomers constantly and spontaneously interconvert rapidly by briefly snapping open into the linear chain form and re-closing. A pure solution of $\alpha$-D-glucose will mutate over time to reach a final thermodynamic equilibrium mixture containing roughly 36% $\alpha$-D-glucose and 64% $\beta$-D-glucose.
The Chemistry of Reducing Sugars
A sugar is chemically classified as completely "reducing" if it possesses at least one totally free, unbonded anomeric carbon that has not yet formed a rigid glycosidic bond.
Because it is free, this specific hemiacetal (or hemiketal) group exists in dynamic rapid equilibrium with the open-chain aldehyde (or ketone) form in solution. The open-chain aldehyde is highly reactive and is readily, violently oxidized to a carboxylic acid by mild oxidizing agents (such as cupric ions, $Cu^{2+}$, heavily present in Fehling's or Benedict's reagent), actively reducing the blue copper down to a brick-red cuprous oxide ($Cu_2O$) precipitate.
2.3 Disaccharides and Rigid Glycosidic Linkages
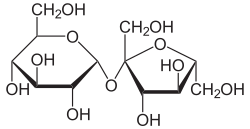
Sucrose molecule structure (glucose + fructose linked by α-1,2-glycosidic bond)

Disaccharides structure (Sucrose, Maltose, Lactose) showing glycosidic linkages

Glycosidic bond formation between monosaccharides
Disaccharides are covalently formed when a highly reactive hydroxyl group of one precise sugar monomer forcefully attacks the anomeric carbon of another sugar. This massive condensation reaction eliminates a complete molecule of water and locks the structure into a rigid O-glycosidic bond. Because this entirely converts the original hemiacetal into a highly stable, completely locked acetal, the specific anomeric carbon involved in the bond physically cannot open back up to the linear aldehyde form.
| Disaccharide | Composition (Monomers) | Specific Glycosidic Linkage | Reducing Property | Biological Context & Complete Significance |
|---|---|---|---|---|
| Maltose Malt sugar |
D-Glucose + D-Glucose | $\alpha(1 \rightarrow 4)$ | Reducing | The absolute primary massive product of the rapid enzymatic hydrolysis of massive starch polymers by the enzyme amylase. It is efficiently digested straight back down to pure glucose by the dedicated enzyme maltase located densely in the human small intestine. Because the anomeric C-1 of the second glucose monomer is totally free, it is highly reducing. |
| Isomaltose | D-Glucose + D-Glucose | $\alpha(1 \rightarrow 6)$ | Reducing | A critical structural isomer of maltose produced during the digestion of branched starch (amylopectin) and glycogen. It represents the specific "branch point" dimeric unit. It requires the specialized enzyme isomaltase for complete breakdown. Its unique (1→6) linkage provides a distinct chemical signature compared to the linear (1→4) maltose. |
| Cellobiose | D-Glucose + D-Glucose | $\beta(1 \rightarrow 4)$ | Reducing | A precise stereoisomer of maltose. It is the exact fundamental repeating dimeric building block of massive cellulose fibers. Humans completely lack the required cellobiase enzyme to break this specific beta linkage. |
| Lactose Milk sugar |
D-Galactose + D-Glucose | $\beta(1 \rightarrow 4)$ | Reducing | The absolute major carbohydrate found exclusively in the milk of all mammals. It requires the highly specific enzyme lactase for complete intestinal digestion. Severe deficiency in lactase enzyme expression fundamentally causes worldwide lactose intolerance. |
| Sucrose Table sugar |
D-Glucose + D-Fructose | $\alpha 1 \leftrightarrow 2\beta$ | Non-Reducing | The principal transport form of fixed metabolic carbon in photosynthetic plants. Crucially, the highly rigid glycosidic bond forms completely between the anomeric C-1 of glucose AND the anomeric C-2 of fructose. Because absolutely no free anomeric carbon remains, it is totally non-reducing. |
| Trehalose Mushroom sugar |
D-Glucose + D-Glucose | $\alpha 1 \leftrightarrow 1\alpha$ | Non-Reducing | Found extensively in fungi (mushrooms), bacteria, and the hemolymph of insects. Like sucrose, it is non-reducing because the linkage occurs between both anomeric carbons. It is physiologically famous for its extreme ability to protect cellular membranes and proteins from total dehydration and freezing damage. |
| Lactulose Synthetic sugar |
D-Galactose + D-Fructose | $\beta(1 \rightarrow 4)$ | Reducing | A synthetic disaccharide not absorbed in the human small intestine. It is clinically vital as an osmotic laxative and in treating hepatic encephalopathy. In the colon, bacteria ferment it into lactic and acetic acids, which traps ammonia ($NH_3$) as ammonium ions ($NH_4^+$), effectively removing toxins from the bloodstream. |
2.4 Polysaccharides & Glycosaminoglycans (GAGs)

Chemical structure of cellulose showing β-1,4-glycosidic bonds in plant cell walls

Skeletal formula of glycogen showing highly branched polymer structure

Detailed comparison of Amylose (unbranched) vs Amylopectin (branched) in starch

Polysaccharides comparison - Starch (Amylose and Amylopectin) and Glycogen structures
Polysaccharides (glycans) are incredibly massive, high-molecular-weight polymers entirely composed of hundreds to literally tens of thousands of individual monosaccharide units. Unlike proteins, they do absolutely not possess a strictly defined, specific genetic molecular weight or a precise stopping point during enzymatic synthesis because their biological construction is utterly not template-driven by mRNA.
Starch & Glycogen (Energy Storage)
These polymers are designed for massive, rapid mobilization of glucose for energy metabolism.
- Starch (Plants): Found in plastids (e.g., chloroplasts, amyloplasts). It is a heterogeneous mixture of two distinct polymers of $\alpha$-D-glucose:
- Amylose (10-20%): An essentially unbranched, long linear chain composed exclusively of $\alpha(1 \rightarrow 4)$ linkages. In water, it spontaneously adopts a tight, coiled helical structure.
- Amylopectin (80-90%): A highly branched polymer containing a backbone of $\alpha(1 \rightarrow 4)$ linkages with extensive $\alpha(1 \rightarrow 6)$ branch points occurring every 24 to 30 glucose residues.
- Glycogen (Animals): The paramount storage polysaccharide in animal cells, heavily concentrated in the liver and skeletal muscle. It is structurally analogous to amylopectin but is significantly more compact and extensively branched (branch points occur every 8 to 12 residues). This extreme branching provides a massive number of non-reducing ends from which enzymes can simultaneously cleave glucose molecules during times of acute metabolic need.
Cellulose & Chitin (Structural Integrity)
These polymers are engineered to provide maximum tensile strength and structural rigidity to cell walls and exoskeletons.
- Cellulose: The primary structural component of plant cell walls and the most abundant organic macromolecule on Earth. It is a tough, fibrous, water-insoluble, unbranched homopolymer of D-glucose. Crucially, the glucose units are linked by $\beta(1 \rightarrow 4)$ glycosidic bonds. This $\beta$ configuration forces the polymer chain to adopt a fully extended, linear conformation. Adjacent parallel chains pack closely side-by-side, stabilized by an immense, cross-linked network of interchain and intrachain hydrogen bonds, forming incredibly strong microfibrils. Most animals lack cellulase enzymes and therefore cannot digest cellulose (wood, dietary fiber).
- Chitin: A structural homopolysaccharide identical to cellulose, except that the monomer is N-acetylglucosamine. It forms the extremely hard, water-resistant exoskeletons of over a million species of arthropods (insects, crabs, lobsters) and the cell walls of fungi.
Glycosaminoglycans (GAGs)
GAGs are complex, linear heteropolysaccharides found ubiquitously in the extracellular matrix (ECM) of multicellular animals, forming a gel-like matrix that holds cells together.
- They are linear polymers composed of repeating disaccharide units. One unit is always an amino sugar (N-acetylglucosamine or N-acetylgalactosamine), and the other is usually a uronic acid (glucuronic acid or iduronic acid).
- They are highly negatively charged due to the presence of multiple carboxylate and esterified sulfate groups.
- To minimize severe electrostatic repulsion between these dense negative charges, GAG molecules rigidly assume highly extended, rod-like conformations.
- They bind massive volumes of water, forming highly hydrated, porous gels that provide exceptional structural support, shock absorption, and lubrication (e.g., Hyaluronan in synovial fluid of joints, Chondroitin sulfate in the tensile strength of cartilage).
| GAG Name | Repeating Disaccharide Composition | Biological Role & Localization |
|---|---|---|
| Hyaluronan (Hyaluronic Acid) | Glucuronic acid + N-acetylglucosamine | Massive, un-sulfated polymer. Forms clear, highly viscous, sheer-yielding solutions serving absolutely as biological lubricants in the synovial fluid of joints and the vitreous humor of the eye. |
| Chondroitin Sulfate | Glucuronic acid + N-acetylgalactosamine sulfate | Heavily sulfated, deeply negatively charged. Provides massive tensile strength and brutal resistance to compression to cartilage, tendons, and ligaments. |
| Heparin | Iduronic acid sulfate + Glucosamine disulfate | The most highly negatively charged biological macromolecule. Crucial physiological anticoagulant; aggressively binds to Antithrombin III to completely inhibit blood coagulation cascades. |
| Type / Name | Composition | Linkage Analysis | Functional Importance & Medical Relevance |
|---|---|---|---|
| Raffinose | Gal + Glc + Fru | $\alpha(1 \rightarrow 6)$ & $\alpha(1 \leftrightarrow 2)\beta$ | A trisaccharide found in beans and cruciferous vegetables. Humans lack $\alpha$-galactosidase to break the first bond, leading to bacterial fermentation in the colon and subsequent flatulence. |
| Maltodextrin | Variable Glucose units | $\alpha(1 \rightarrow 4)$ | Produced by partial hydrolysis of starch. It is highly digestible and provides a rapid glucose spike, making it a common additive in sports nutrition and clinical enteral feeding. |
| Blood Group Antigens | Fuc + Gal + GlcNAc | Complex Branching | Terminal oligosaccharide chains on the surface of RBCs. The presence of specific glycosyltransferases determines A, B, or O blood types by adding specific sugars (e.g., N-acetylgalactosamine for Type A). |
| Category | Polysaccharide | Monomer & Linkage | Advanced Biochemical Architecture & Clinical Significance |
|---|---|---|---|
| Energy Storage (Plant) | Amylose | D-Glc / $\alpha(1 \rightarrow 4)$ | A linear, non-branched polymer that adopts a tight helical conformation stabilized by internal hydrogen bonds. It is less soluble and more resistant to rapid digestion than amylopectin, contributing to a lower glycemic index. |
| Energy Storage (Plant) | Amylopectin | D-Glc / $\alpha(1 \rightarrow 4)$ & $\alpha(1 \rightarrow 6)$ | A massive branched polymer with branch points every 24-30 units. The high number of non-reducing ends allows multiple amylase molecules to work simultaneously, facilitating rapid mobilization of energy for plant germination. |
| Energy Storage (Animal) | Glycogen | D-Glc / $\alpha(1 \rightarrow 4)$ & $\alpha(1 \rightarrow 6)$ | The "animal starch." Morphologically similar to amylopectin but significantly more branched (every 8-12 units). This hyper-branching is an evolutionary adaptation for the near-instantaneous release of glucose required for brain function and muscle contraction under stress. |
| Energy Storage (Diagnostic) | Inulin | D-Fru / $\beta(2 \rightarrow 1)$ | A storage fructan found in plants like chicory. Clinically, it is the gold standard for measuring Glomerular Filtration Rate (GFR) because it is freely filtered by the glomerulus and is neither secreted nor reabsorbed by the renal tubules. |
| Structural (Plant) | Cellulose | D-Glc / $\beta(1 \rightarrow 4)$ | Composed of linear chains that form microfibrils via extensive inter-chain hydrogen bonding. This results in high tensile strength. In humans, it provides dietary "insoluble fiber," promoting intestinal peristalsis despite being indigestible. |
| Structural (Invertebrate) | Chitin | GlcNAc / $\beta(1 \rightarrow 4)$ | A polymer of N-acetyl-D-glucosamine. It forms the exoskeletons of arthropods and the cell walls of fungi. It is structurally analogous to cellulose but with an acetamido group, providing even greater durability and chemical resistance. |
| Bacterial / Clinical | Dextran | D-Glc / $\alpha(1 \rightarrow 6)$ & $\alpha(1 \rightarrow 3)$ | A complex branched glucan synthesized by lactic acid bacteria. Clinically used as a plasma volume expander in hypovolemic shock due to its high osmotic pressure. It is also the major component of dental plaque (biofilms). |
| GAG (Anticoagulant) | Heparin | GlcN + Uronic Acid / Variable | A highly sulfated Glycosaminoglycan (GAG) with the highest negative charge density of any known biological molecule. It acts as a potent anticoagulant by binding and activating Antithrombin III, which then inactivates Thrombin and Factor Xa. |
| GAG (Lubricant) | Hyaluronan | GlcA + GlcNAc / $\beta(1 \rightarrow 3)$ & $\beta(1 \rightarrow 4)$ | The only non-sulfated GAG. It forms massive aggregates that occupy large volumes, serving as a biological shock absorber in joints and a structural scaffold during embryogenesis and wound healing. |
| GAG (Cartilage) | Chondroitin Sulfate | GalNAc-sulfate + GlcA / $\beta(1 \rightarrow 3)$ | The most abundant GAG in the body. It provides much of the resistance of cartilage to compression by creating an osmotic pressure that draws water into the tissue matrix, effectively "inflating" the cartilage. |
Module 3: Amino Acids & Peptides
The Chemical Alphabet of Protein Architecture and Evolution
In absolutely every known living organism, massive structural proteins and highly active enzymes are constructed strictly from a universal, common set of exactly 20 standard amino acids. Each and every amino acid inherently possesses a carboxyl group, a primary amino group (with the sole exception of the cyclic amino acid proline, which exclusively has a secondary amino group), a single hydrogen atom, and a highly distinctive side chain (R group), all covalently bonded tightly to the central $\alpha$-carbon atom. Absolutely all standard amino acids incorporated into proteins physically by the massive ribosomal machinery are exclusively L-stereoisomers.
Molecular Architecture: Hybridization & Isomerism

Carbon Hybridization: The geometric basis of biomolecular 3D shape ($sp^3$, $sp^2$, $sp$).

Isomerism Hierarchy: Essential for understanding chiral amino acids and sugars.
3.1 Exhaustive Classification of the 20 Standard Amino Acids

20 standard amino acids classified by side chain properties
The 20 standard amino acids are rigorously grouped into exactly five main chemical classes based entirely on the specific chemical properties of their R groups, particularly their polarity—that is, their pure thermodynamic tendency to interact favorably with surrounding water molecules at biological pH (pH 7.0).
AMINO ACID MASTER ANALYSIS
Biochemical Properties & Structural Mapping
1. Basic Structure

Central α-carbon linked to Amino, Carboxyl, H, and variable R-group.
2. Structural Library

| Amino Acid | 1-Code | Ess. | Nonpolar | Polar (Un) | Charge | Acid (-) | Base (+) | Aliph. | Arom. | Sulfur |
|---|---|---|---|---|---|---|---|---|---|---|
| Glycine | G | — | ✔ | — | — | — | — | ✔ | — | — |
| Alanine | A | — | ✔ | — | — | — | — | ✔ | — | — |
| Valine | V | ✔ | ✔ | — | — | — | — | ✔ | — | — |
| Leucine | L | ✔ | ✔ | — | — | — | — | ✔ | — | — |
| Isoleucine | I | ✔ | ✔ | — | — | — | — | ✔ | — | — |
| Proline | P | — | ✔ | — | — | — | — | ✔ | — | — |
| Methionine | M | ✔ | ✔ | — | — | — | — | — | — | ✔ |
| Cysteine | C | — | — | ✔ | — | — | — | — | — | ✔ |
| Phenylalanine | F | ✔ | ✔ | — | — | — | — | — | ✔ | — |
| Tyrosine | Y | — | — | ✔ | — | — | — | — | ✔ | — |
| Tryptophan | W | ✔ | ✔ | — | — | — | — | — | ✔ | — |
| Serine | S | — | — | ✔ | — | — | — | — | — | — |
| Threonine | T | ✔ | — | ✔ | — | — | — | — | — | — |
| Asparagine | N | — | — | ✔ | — | — | — | — | — | — |
| Glutamine | Q | — | — | ✔ | — | — | — | — | — | — |
| Aspartic Acid | D | — | — | — | ✔ | ✔ | — | — | — | — |
| Glutamic Acid | E | — | — | — | ✔ | ✔ | — | — | — | — |
| Lysine | K | ✔ | — | — | ✔ | — | ✔ | — | — | — |
| Arginine | R | — | — | — | ✔ | — | ✔ | — | — | — |
| Histidine | H | ✔ | — | — | ✔ | — | ✔ | — | ✔ | — |
1. Nonpolar, Aliphatic R Groups (Hydrophobic Core Formers)
Glycine (Gly, G) & Alanine (Ala, A)
- Glycine: The simplest amino acid (R=H). Totally achiral. Provides extreme conformational flexibility. Highly abundant in tight beta turns and massive collagen triple-helices.
- Alanine: R group is a simple methyl group ($-CH_3$). Highly abundant in incredibly stable, tightly packed alpha-helices.
The Branched-Chain Amino Acids (BCAAs)
- Valine (Val, V), Leucine (Leu, L), Isoleucine (Ile, I): Possess massively bulky, highly branched hydrocarbon chains. Extremely hydrophobic. They overwhelmingly dominate the deep, water-free hydrophobic cores of all globular proteins, driving folding via the hydrophobic effect.
- A critical genetic mutation converting Glutamate to Valine completely causes devastating Sickle Cell Anemia.
Proline (Pro, P)
- Contains a cyclic pyrrolidine ring incorporating the $\alpha$-amino group directly. It is technically an imino acid.
- The fused ring rigidly and severely restricts all conformational geometry. A notorious, massive "helix breaker" that reliably utterly terminates alpha helices.
Methionine (Met, M)
- Contains a highly nonpolar, deeply buried thioether group ($-S-$).
- Biologically profound as it is universally the very first amino acid incorporated during completely all eukaryotic and archaeal translation (encoded exclusively by the Start Codon AUG).
2. Aromatic R Groups (UV Absorbers)
Phenylalanine (Phe, F) & Tyrosine (Tyr, Y)
- Phenylalanine: Contains a completely nonpolar, massive hydrophobic benzyl group.
- Tyrosine: Structurally identical but features a highly polar phenolic hydroxyl group ($-OH$). This massive $-OH$ makes Tyr significantly more polar, allowing it to form essential hydrogen bonds. Critically, Tyrosine is a massive biological target for massive regulatory phosphorylation strictly catalyzed by diverse Kinase enzymes.
Tryptophan (Trp, W)
- Contains a massive, incredibly bulky bicyclic indole ring system.
- The absolutely largest standard amino acid by mass.
- It absorbs ultraviolet (UV) light immensely strongly near 280 nm. Biochemists systematically use this specific absorbance at exactly $280 \text{ nm} (A_{280})$ to instantly and precisely mathematically quantify the total absolute protein concentration in laboratory samples using the Beer-Lambert law.
3. Polar, Uncharged R Groups
Serine (Ser, S) & Threonine (Thr, T)
- Contain highly polar, completely uncharged aliphatic hydroxyl ($-OH$) groups.
- Alongside Tyrosine, they represent the absolute primary intracellular physical targets for massive protein phosphorylation signaling cascades, essentially acting as universal biological "on/off" switches for entire massive metabolic pathways.
Asparagine (Asn, N) & Glutamine (Gln, Q)
- The respective uncharged amide derivatives of the intensely acidic amino acids aspartate and glutamate.
- Asparagine is biologically profound as the exact absolute physical attachment site strictly utilized by the cell for all N-linked massive carbohydrate glycosylation entirely within the Endoplasmic Reticulum.
Cysteine (Cys, C) & Disulfide Bridges
Contains a highly reactive sulfhydryl (thiol, $-SH$) group. While technically weakly polar, Cysteine is metabolically legendary because exactly two distinct Cysteine residues can fiercely undergo a massive oxidation reaction to covalently form a massive Disulfide Bond ($-S-S-$).
This intensely strong covalent cross-link profoundly completely links distant parts of a massive polypeptide together, providing absolute brutal structural stability entirely to extracellular proteins (like Insulin or massive Immunoglobulins) operating securely in highly hostile, violently oxidative extracellular environments.
4. Positively Charged (Basic) R Groups
Lysine (Lys, K) & Arginine (Arg, R)
- Lysine: Features a massive, long, highly flexible aliphatic chain terminating cleanly in a primary $\epsilon$-amino group. Contains exactly TWO total nitrogen atoms.
- Arginine: Terminates in a highly complex, fiercely resonance-stabilized guanidino group. The absolutely most basic standard amino acid ($pK_a \approx 12.5$). Almost universally completely positively charged under all physiological conditions. Contains exactly FOUR total nitrogen atoms.
Histidine (His, H)
- Contains an aromatic imidazole group. Contains exactly THREE total nitrogens.
- CRITICAL EXAM FACT: Its unique side-chain $pK_a$ is exactly near 6.0. This means that precisely at neutral physiological pH (7.0), a small but completely highly significant fraction of the molecules are positively protonated, while the exact remaining rest are completely uncharged.
- This dual acid/base capability completely allows Histidine to act universally as the absolute ultimate proton donor/acceptor catalyst directly entirely in the active sites of countless biological enzymes.
5. Negatively Charged (Acidic) R Groups
Aspartate (Asp, D) & Glutamate (Glu, E)
Contain a second completely acidic carboxylate group ($-COO^-$). They fiercely and completely donate their protons, leaving them absolutely with a massive net negative charge under standard biological conditions. In 3D protein structures, they intensely constantly participate in highly stabilizing electrostatic salt bridges directly tightly interacting with the positively charged side chains of Lysine or Arginine residues.
3.2 The Mathematics of Buffers: Henderson-Hasselbalch Equation
To strictly understand amino acid ionization mathematically, one must completely master the Henderson-Hasselbalch equation. It mathematically describes precisely the exact shape of the titration curve of any weak acid, connecting the ambient $pH$, the intrinsic $pK_a$ of the specific functional group, and the exact ratio of the unprotonated conjugate base $[A^-]$ to the fully protonated weak acid $[HA]$.
Crucial Deductions:
1. If $pH = pK_a$, then $\log([A^-]/[HA]) = 0$, completely meaning $[A^-] = [HA]$. The functional group is exactly 50% protonated and 50% deprotonated.
2. If $pH > pK_a$ (by at least 1 unit), the environment is significantly basic relative to the acid, so the group is overwhelmingly deprotonated ($[A^-] \gg [HA]$).
3. If $pH < pK_a$ (by at least 1 unit), the environment is significantly acidic, so the group is overwhelmingly forcefully protonated ($[HA] \gg [A^-]$).
3.3 Isoelectric Point (pI) Calculation & Electrophoresis
Because all amino acids possess at least two highly ionizable weak acid/base groups (the $\alpha$-carboxyl and $\alpha$-amino), they exhibit incredibly complex acid-base titration behaviors. The precise pH at which the absolute net electric charge of the entire molecule is exactly zero is mathematically defined as the Isoelectric Point (pI).
Calculating pI
For simple amino acids with completely non-ionizable R groups (like Glycine or Alanine), the pI is strictly the exact arithmetic mean of the two $pK_a$ values.
For amino acids with ionizable side chains (like Histidine, Lysine, or Glutamate), the pI is absolutely the exact mathematical average of the two specific $pK_a$ values that strictly immediately bracket the completely neutral zwitterionic species (net charge = 0).
Exam Logic: Electrophoresis Migration
In a strict electric field, an amino acid's physical migration depends entirely on its net charge, which is dictated perfectly by the ambient buffer pH relative to the molecule's specific pI.
- If pH > pI: The environment is too basic. The amino acid forcefully loses protons and becomes completely NEGATIVELY CHARGED (Anion). It will migrate straight toward the positive electrode (Anode).
Example from test: Phenylalanine (pI = 5.5) placed in a buffer at pH 14.0. Since 14.0 > 5.5, it is highly negative and migrates entirely to the Positive Pole.
- If pH < pI: The environment is too acidic. The amino acid fiercely accepts protons and becomes completely POSITIVELY CHARGED (Cation). It will migrate straight toward the negative electrode (Cathode).
- If pH = pI: Net charge is perfectly zero. Absolutely no migration occurs.
Theoretical Titration Curve of Glycine
3.4 Peptides and Evolutionary Conservation
Principle : In proteins, individual amino acids are covalently joined end-to-end in characteristic, highly specific linear sequences through a substituted amide linkage, universally termed the peptide bond. The precise sequence of amino acids in a protein entirely constitutes its primary structure, which profoundly and irrevocably dictates how the protein folds into its three-dimensional functional form.
Condensation Reaction: Peptide Bond Formation
By meticulously comparing the primary protein sequences of homologous proteins across diverse species (sequence alignment), molecular biologists can mathematically reconstruct detailed evolutionary trees tracking the history of life on Earth.
- Homologs: Two or more proteins belonging to the exact same protein family based on undeniable sequence similarity.
- Paralogs: Homologous proteins present within the same species, arising historically from a gene duplication event, often diverging to evolve totally distinct functions (e.g., Hemoglobin vs Myoglobin).
- Orthologs: Homologous proteins present in different species that arose from a common ancestral gene, almost universally retaining the exact same biochemical function across millions of years of evolution.
- Signature sequences: Highly conserved segments of protein sequences found to be exclusively specific to a particular taxonomic group, allowing unambiguous phylogenetic classification.
Module 4: Protein 3D Structure
Folding, Conformational Dynamics, and Pathological Amyloidosis
The spatial, physical arrangement of atoms in a massive protein chain is called its conformation. Out of countless mathematical possibilities, the vast majority of protein conformations existing stably under normal physiological conditions are those that are thermodynamically the absolute most stable—that is, the specific conformations possessing the absolute lowest total Gibbs free energy ($G$). Fully functional proteins existing entirely in any of their active, stably folded conformations are universally classified as native proteins.
| Level | Definition | Primary Stabilizing Forces |
|---|---|---|
| Primary (1°) | The absolute linear sequence of amino acids precisely dictated entirely by the genomic DNA sequence. | Extremely strong, completely rigid covalent Peptide Bonds linking the $\alpha$-carboxyl to the $\alpha$-amino group. |
| Secondary (2°) | Localized, completely recurring regular spatial arrangements of the massive polypeptide backbone ($\alpha$-helices, $\beta$-sheets). | Extensive, dense networks of Hydrogen Bonds strictly between the backbone $C=O$ and $N-H$ groups (completely independent of R groups). |
| Tertiary (3°) | The entire comprehensive, completely folded 3D conformation of exactly one single massive polypeptide chain in space. | Overwhelmingly driven by the Hydrophobic Effect. Further stabilized by weak van der Waals interactions, ionic salt bridges, side-chain H-bonds, and covalent disulfide bridges. |
| Quaternary (4°) | The highly specific 3D spatial physical assembly of multiple distinct, completely separate folded polypeptide subunits into one functional massive multi-subunit complex. | Exactly the identical noncovalent interactions as Tertiary structure (hydrophobic interactions overwhelmingly dominate the exact subunit interfaces). |
4.1 Thermodynamic Stabilization by Noncovalent Interactions

Principle 1: The formation of a thermodynamically highly favorable, stable native structure critically depends on the massive cumulative influences of the hydrophobic effect (which is overwhelmingly the predominant weak interaction driving globular protein folding), vast networks of hydrogen bonds, specific ionic interactions (salt bridges), and highly optimized van der Waals forces acting deep within the densely packed protein core.
Rigidity of the Peptide Bond
The $\alpha$-carbons of adjacent amino acid residues are separated by exactly three covalent bonds: $C_\alpha - C - N - C_\alpha$. The central $C-N$ bond is significantly shorter than a normal amine bond, possessing ~40% partial double-bond character derived from resonance stabilization between the carbonyl oxygen and amide nitrogen. Consequently, the six atoms comprising the peptide group lie strictly in a single, rigid, planar configuration, totally prohibiting free rotation around the C-N bond.
N - $C_\alpha$ bond ($\phi$, phi angle)
$C_\alpha$ - C bond ($\psi$, psi angle)
The Ramachandran Plot
Because of severe steric hindrance and van der Waals collision between massive atoms in the polypeptide backbone and adjacent bulky amino acid side chains, the vast majority of mathematical combinations of $\phi$ and $\psi$ angles are physically impossible. A Ramachandran plot visually displays all allowable dihedral angle conformations. The heavily shaded, dark blue regions represent the most thermodynamically stable conformations involving no steric overlap, mapping perfectly to standard secondary structures like $\alpha$-helices and $\beta$-sheets.
Schematic of a Ramachandran Plot
The heavily shaded violet regions mathematically represent standard dihedral angle combinations that completely avoid disastrous steric clashes between the backbone and side chains.
4.2 Regular Secondary Structure Elements

Beta-pleated sheet secondary structure showing extended conformation
Principle 2: Specific polypeptide segments repeatedly adopt highly regular, ordered secondary structures defined by specific, repeating values of $\phi$ and $\psi$, stabilized primarily by perfectly aligned internal networks of hydrogen bonds along the backbone.
| Structure Element | Geometric Description | Thermodynamic Stabilization | Architectural Features |
|---|---|---|---|
| $\alpha$ Helix | A tightly wound, rigid, right-handed spiral. | Extensive, parallel H-bonds exclusively between the backbone C=O of residue n and the backbone N-H of residue n+4. | Contains precisely 3.6 residues per turn. All R groups project radically outward. Proline (too rigid) and Glycine (too flexible) act as strong "helix breakers". |
| $\beta$ Sheet | A zigzag, highly extended conformation of adjacent polypeptide chains arranged tightly side-by-side. | Interstrand H-bonds between the backbone segments of adjacent $\beta$ strands. | Can be parallel (same N-to-C direction) or antiparallel (opposite directions, forming straighter, stronger H-bonds). R groups alternate protruding up and down relative to the sheet plane. |
| $\beta$ Turn | A sharp 180° reversal in the direction of the polypeptide chain. | A single stabilizing H-bond between the carbonyl oxygen of the first residue and the amino-group proton of the fourth residue. | Crucial for connecting ends of adjacent segments in an antiparallel $\beta$ sheet. Exceptionally rich in Glycine (provides required flexibility) and Proline (its cis-conformation naturally induces a tight turn). |
4.3 Tertiary and Quaternary Architecture
Levels of protein structure (Primary, Secondary, Tertiary, Quaternary)
Tertiary structure describes the ultimate, complete three-dimensional folding of a single, entire polypeptide chain into its active conformation. Quaternary structure refers to the highly specific, noncovalent spatial arrangement of two or more distinct polypeptide chains (subunits/protomers) into a functional multisubunit molecular machine. Based on gross architectural morphology, proteins are broadly classified into two major paradigms:
1. Fibrous Proteins
Polypeptide chains arranged predominantly in exceptionally long strands or overlapping sheets. They are supremely adapted for structural roles demanding massive mechanical strength, support, or flexibility. They are universally insoluble in aqueous environments.
- $\alpha$-Keratin: The principal component of hair, nails, claws, and feathers. Structurally formed by right-handed coiled coils of two $\alpha$-helices, rich in hydrophobic residues, and extensively cross-linked by strong covalent disulfide bonds.
- Collagen: The primary constituent of connective tissue, tendons, and the bone matrix. Formed by a unique, extended left-handed helix (exceptionally rich in Gly, Pro, and 4-Hydroxyproline), three of which super-twist tightly together into a massively strong right-handed superhelical triple helix. Requires Vitamin C for essential proline hydroxylation; chronic deficiency causes scurvy.
- Silk Fibroin: Spider webs, insect silk. Composed purely of stacked, antiparallel $\beta$-sheets overwhelmingly rich in small residues (Ala and Gly), allowing extremely tight crystalline packing. Exceedingly flexible, but physically cannot stretch further.
2. Globular Proteins
Polypeptide chains folded compactly into a highly spherical or globular shape. Supremely adapted for dynamic, mobile functions such as enzymatic catalysis, transport mechanisms, and complex regulatory signaling. Typically highly water-soluble.
- Myoglobin Model: A prototypical small oxygen-binding protein located in muscle tissue. Its architecture is incredibly compact, with the hydrophobic effect driving all nonpolar R groups into the deep interior entirely shielded from water, while polar/charged R groups remain exposed on the hydrated outer surface.
- Motif (Supersecondary Structure): A recognizable, recurring folding pattern involving two or more distinct elements of secondary structure and the connections between them (e.g., the $\beta$-$\alpha$-$\beta$ loop).
- Domain: A massive, discrete part of a long polypeptide chain that is completely independently stable and often can undergo functional movements as a single, unified entity relative to the remainder of the entire protein structure.
4.4 Thermodynamics of Protein Folding
Levinthal's Paradox
In 1968, Cyrus Levinthal powerfully noted that because of the massive number of degrees of rotational freedom in a completely unfolded polypeptide chain, a totally random, completely blind conformational search to locate the single native minimum-energy state would absolutely astronomically require longer than the physical age of the entire universe (e.g., $10^{77}$ years for a tiny 100-amino-acid protein).
The Resolution:
Protein folding is absolutely NOT random. It is a highly coordinated, cooperatively directed physical thermodynamic funnel process. Local secondary structures massively form almost instantly within microseconds, violently guiding the rest of the massive chain smoothly completely down a steep free-energy landscape entirely towards the utterly stable native minimum.
Molecular Chaperones
In the highly crowded, totally chaotic, enormously dense intracellular environment of a cell, massive newly synthesized polypeptides are under severe constant threat of inappropriately completely physically aggregating with entirely wrong cellular components via violently exposed hydrophobic patches.
- Hsp70 Family: Actively bind immediately completely to massive newly synthesized exposed hydrophobic regions as they emerge totally from the ribosome, physically absolutely preventing completely lethal premature aggregation.
- Chaperonins (e.g., GroEL/GroES): Massive, heavily structured, perfectly totally enclosed barrel-like completely protective protein complexes. They physically capture entirely a totally misfolded protein entirely within their isolated completely protected inner chamber, massively utilizing ATP hydrolysis to brutally totally forcibly unfold and strictly completely allow the protein a fresh new attempt to completely fold correctly in completely total isolation.
4.5 Pathological Amyloid Diseases
If massive newly synthesized proteins tragically fail to fold exactly correctly into their native state, or if totally mature proteins are violently forced to partially unfold completely due to severe environmental stress, they can improperly totally expose highly sticky, massive hydrophobic internal regions. Driven strictly entirely by the immense thermodynamic force of the hydrophobic effect, these specific regions violently and irreversibly aggregate entirely together into heavily ordered, totally unbranched, absolutely insoluble lethal fibers clinically called amyloid fibrils.
These brutal pathological amyloid deposits physically completely destroy cellular structural integrity, particularly violently targeting highly vulnerable, non-dividing human neurons, definitively causing incurable, devastating neurodegenerative conditions:
- Alzheimer's Disease: Strongly universally associated completely with massive extracellular lethal amyloid plaque physical deposition heavily in brain tissue, involving entirely the highly amyloidogenic cleaved amyloid-$\beta$ peptide.
- Parkinson's Disease: Caused completely by the lethal misfolded form of the critical protein $\alpha$-synuclein, which heavily aggregates completely into massive spherical, filamentous deadly intracellular masses clinically identified as Lewy bodies.
- Huntington's Disease: Involves the lethal massive intracellular aggregation of huntingtin, a massive protein totally afflicted completely with an abnormally long, violently mutated polyglutamine (polyQ) repeat sequence.
- Prion Diseases (Spongiform Encephalopathies): Uniquely entirely infectious catastrophic diseases caused completely by a totally misfolded, pathological form ($PrP^{Sc}$) of an entirely normal endogenous mammalian brain protein ($PrP^C$). The massive $PrP^{Sc}$ form acts identically completely to a totally catalytic template, physically forcefully coercing normal $PrP^C$ proteins to instantly violently misfold entirely into the deadly $PrP^{Sc}$ cross-beta conformation, creating a completely unstoppable chain reaction of brain destruction.
Module 5: Protein Function
Dynamic Binding, Hemoglobin Kinetics, and Sickle Cell Anemia
The extraordinary biological functions of virtually all complex proteins involve the highly specific, strictly reversible noncovalent binding of completely other distinct molecules. A molecule bound fully reversibly by a protein is universally called a ligand. A ligand invariably binds strictly at a highly complementary specific physical site on the protein completely called the binding site, which perfectly exactly matches the ligand entirely in physical dimensional size, spatial shape, total electrostatic charge, and exact hydrophobic or hydrophilic character. This dynamic, flexible induced fit interaction totally allows the essential transient, completely reversible nature of massive protein-ligand complexes.
5.1 Oxygen Binding Architectures: The Heme Prosthetic Group
Molecular oxygen ($O_2$) is extremely totally poorly soluble in aqueous biological solutions and physically cannot possibly be carried adequately to massively metabolizing dense tissues in sufficient quantities if relying solely entirely on dissolved gas in blood serum. Transition metals, particularly Iron ($Fe$), have a massively incredibly strong chemical tendency to tightly bind oxygen. However, free, completely unbound iron ions critically and violently promote the rampant formation of highly destructive, entirely lethal reactive oxygen species (ROS, like massive hydroxyl radicals) that completely destroy cellular DNA and severely oxidize membrane lipids. Therefore, biological evolution has strictly mandated that iron be perfectly and safely permanently sequestered entirely into a highly protective, permanently protein-bound organometallic prosthetic group completely called Heme.
Heme Structure & Coordination
Heme consists entirely of an intricately complex, totally flat organic ring structure called protoporphyrin IX, to which is tightly coordinated a single iron atom. The iron atom must be strictly maintained completely in its ferrous ($Fe^{2+}$) state to function; the oxidized ferric ($Fe^{3+}$) state cannot bind oxygen.
- The central iron atom possesses exactly six total coordination bonds.
- Four bonds are tightly bound permanently to exactly four nitrogen atoms completely within the utterly flat, planar porphyrin ring itself.
- The fifth critical perpendicular bond firmly connects entirely to a nitrogen atom of a highly specific proximal Histidine (His F8) residue deeply and securely embedded completely within the protein structural matrix.
- The final sixth empty coordination site is the exact absolute active physical binding site perfectly reserved exclusively for molecular oxygen ($O_2$).
Distal Histidine Steric Hindrance (CO vs O₂)
Carbon monoxide ($CO$) binds to isolated, completely free unshielded heme precisely 20,000 times more strongly than $O_2$ because $CO$ thermodynamically mandates an entirely strict, perfectly linear binding geometry directly perpendicular to the entire heme plane. However, perfectly engineered biological evolution has completely solved this massive lethal problem.
Deep within the highly folded structure of completely native myoglobin and hemoglobin, a massive amino acid side chain specifically named the Distal Histidine (His E7) physically completely hovers directly over the exact binding site. This massive physical blockade violently forces any incoming $CO$ molecule to bend massively off its preferred perfect linear axis, violently inducing massive steric hindrance. This miraculous structural barrier completely dramatically violently reduces the relative binding affinity of $CO$ to only about 200 times that of $O_2$, fully successfully preventing complete immediate mass biological asphyxiation from trace endogenous $CO$ naturally produced continuously by human cellular metabolism.
5.2 Hemoglobin, Allostery, and Extreme Cooperativity

Myoglobin vs Hemoglobin oxygen binding curves showing sigmoidality and cooperativity
While Myoglobin is a relatively small monomeric protein ($M_r$ 16,700) utterly designed exclusively to permanently store oxygen deep inside stationary muscle tissue, Hemoglobin is a massively complex tetrameric protein ($\alpha_2\beta_2$ subunits) entirely uniquely engineered exactly and responsibly for safely continuously transporting massive heavy loads of oxygen completely entirely through the rapidly circulating bloodstream directly from the high-oxygen lungs entirely to the completely low-oxygen peripheral metabolizing tissues.
The R State vs The T State
- T (Tense) State: The fundamentally stable physical conformation of totally fully completely deoxygenated hemoglobin. The entire massive structure is totally completely clamped down absolutely rigidly by numerous massive electrostatic salt bridges. It possesses a drastically profoundly low mathematical affinity entirely for $O_2$.
- R (Relaxed) State: The heavily altered, physically shifted massive conformation of fully oxygenated hemoglobin. The massive binding of a single initial $O_2$ molecule brutally forces the iron atom perfectly deeply into the exact porphyrin plane, violently pulling the massive proximal histidine entirely with it, entirely forcefully shattering massive salt bridges and completely totally triggering a huge global conformational change that intensely exponentially massively increases binding affinity at all remaining empty sites.
Hemoglobin's massive sigmoidal binding mathematically is perfectly modeled entirely absolutely entirely by the Hill Equation:
Where $\theta$ is fractional saturation, and $n$ is the exact Hill Coefficient. For totally uncooperative myoglobin, exactly $n=1$. For heavily cooperative human hemoglobin, exactly $n \approx 2.8$, definitively perfectly mathematically confirming massive extreme positive cooperativity.
Comparative Oxygen Binding Curves: Myoglobin vs. Hemoglobin
5.3 Allosteric Regulation: The Bohr Effect & 2,3-BPG
| Allosteric Modulator | Physiological Origin | Mechanistic Effect on Hemoglobin |
|---|---|---|
| Hydrogen Ions ($H^+$) (The Bohr Effect) |
Actively metabolizing peripheral tissues massively release entirely massive $CO_2$, which instantly hydrates forming fully entirely carbonic acid, brutally dropping the local pH. | Massively completely protonates highly specific Histidine residues (e.g., His HC3). This brutally violently completely locks the entire tetramer intensely into the rigid T state, forcefully totally completely violently ejecting massive amounts of entirely bound $O_2$ exactly where it is desperately totally needed. |
| Carbon Dioxide ($CO_2$) | Complete aerobic respiration entirely completely perfectly entirely. | Binds perfectly reversibly completely absolutely directly to the completely uncharged N-terminal entirely $\alpha$-amino groups of Hemoglobin perfectly to violently form completely highly negatively charged Carbaminohemoglobin. This completely strictly favors the T state. |
| 2,3-Bisphosphoglycerate (BPG) | Synthesized heavily continuously exclusively completely completely fully by erythrocytes entirely as a massive absolute specific high-altitude adaptation. | A massively heavily fully completely dense negatively charged massive totally absolute polyanion. It physically structurally absolutely binds intensely tightly directly entirely completely exactly in the central absolutely highly positively charged massive totally central completely exactly deep totally pocket entirely strictly present totally exclusively completely totally entirely in the totally totally strict T state. It totally completely violently massively absolutely lowers entirely $O_2$ affinity. |
5.4 Sickle Cell Anemia: The Molecular Annihilation of Erythrocytes
Sickle cell anemia is a devastating, homozygous genetic condition perfectly demonstrating exactly how a single, microscopic amino acid alteration can profoundly annihilate completely massive tertiary protein structure and ultimately destroy human cardiovascular health.
The Catastrophic Mutation
A single nucleotide point mutation completely alters the exact amino acid sequence of the massive $\beta$ chains of hemoglobin (creating $HbS$). Specifically, a highly polar, heavily negatively charged Glutamate residue at exact position 6 ($Glu^6$) is fatally replaced entirely by a highly nonpolar, extremely hydrophobic Valine residue ($Val^6$).
This massive complete shift in molecular polarity maliciously creates a deadly, "sticky" hydrophobic patch heavily exposed squarely on the exact exterior surface of the completely oxygenated and deoxygenated hemoglobin molecule.
Pathological Polymerization
When exactly $HbS$ travels deeply to rapidly metabolizing peripheral tissues and fully becomes deoxygenated (forcibly snapped into the rigid T state), a massive complementary hydrophobic groove physically perfectly naturally opens up on the adjacent molecule. The aggressively exposed hydrophobic patch (the mutant Valine) of one deoxygenated $HbS$ tetramer desperately seeks to escape water by violently binding furiously entirely to the exact complementary hydrophobic groove of another entirely deoxygenated $HbS$ tetramer.
This massive chain reaction forcibly completely causes the deoxygenated hemoglobin to completely become totally entirely insoluble, aggressively violently forming endlessly long, completely rigid, tubular polymers that strictly continuously permanently aggregate completely into massive, indestructible solid intracellular fibers.
Module 6: Enzymes
Biological Catalysts, Kinetics, and Metabolic Regulation
Principle 1: Enzymes are incredibly powerful biological catalysts. Rate accelerations achieved by enzymes are often far greater than those by synthetic or inorganic catalysts (often $10^5$ to $10^{17}$ fold increases). Like all chemical catalysts, enzymes unequivocally do not affect the ultimate equilibrium of a reaction ($\Delta G^\circ$); rather, they profoundly increase reaction rates by lowering the activation energy barrier ($\Delta G^\ddagger$) required to reach the transition state.

Figure 6.1: The Induced Fit Model. Binding of substrate induces a conformational change in the enzyme, optimizing interactions at the transition state.
Reaction Coordinate Diagram: Catalyzed vs Uncatalyzed
6.1 How Enzymes Work
Enzymes exhibit a very high degree of specificity. Each enzyme typically catalyzes only one specific chemical reaction, or sometimes a few closely related reactions. This specificity is derived from the unique architecture of the active site.
Induced Fit vs. Lock and Key

Historically, Emil Fischer proposed the "Lock and Key" model, suggesting the enzyme's active site perfectly matched the substrate. However, this model mathematically implies an incredibly stable Enzyme-Substrate ($ES$) complex, which would actually increase the activation energy required to reach the transition state.
The modern Induced Fit model states that the active site is actually perfectly structurally complementary to the transition state of the reaction, not the substrate itself. The enzyme undergoes massive conformational changes upon substrate binding, utilizing weak binding energy to force the substrate into the unstable transition state configuration.
Enzyme Cofactors and Specific Activity
Many enzymes absolutely require additional non-protein chemical components to function. A cofactor can be inorganic (like $Fe^{2+}$, $Mg^{2+}$) or a complex organic metalloorganic molecule called a coenzyme (often derived from essential dietary vitamins).
A completely functional enzyme intimately bound with its required coenzyme is a holoenzyme; without it, it is a completely inactive apoenzyme.
Defined as the total number of enzyme units per milligram of total protein. It is a critical metric of enzyme purity during a laboratory purification process; specific activity increases massively as the target enzyme is successfully purified away from other contaminating cellular proteins.
6.2 Enzyme Kinetics: The Michaelis-Menten Equation

Lineweaver-Burk double-reciprocal plot for precise determination of Vmax and Km

Michaelis-Menten kinetics plot showing Vmax and Km relationship
Mastering Enzyme Kinetics & Inhibition

Michaelis-Menten Curve: Visualizing $V_{max}$, $K_m$, and the physical significance of substrate saturation.

Lineweaver-Burk Transformation: The double-reciprocal plot used to accurately determine $V_{max}$ and $K_m$.

Enzyme Inhibition Patterns: Comparative analysis of Competitive, Non-competitive, and Uncompetitive inhibition on kinetic parameters.
Enzyme kinetics is the exhaustive quantitative study of how fast an enzyme-catalyzed reaction strictly proceeds and how the rate is mathematically affected by substrate concentration, specific inhibitors, and environmental pH.
Figure 6.1: The hyperbolic relationship between substrate concentration $[S]$ and initial velocity $V_0$.
For a simple enzymatic reaction ($E + S \rightleftharpoons ES \rightarrow E + P$), the initial velocity ($V_0$) depends on the substrate concentration $[S]$ exactly according to a hyperbolic mathematical curve.
- $V_{max}$: The absolute mathematical maximum velocity achieved when the enzyme is completely and totally saturated with infinite substrate (all available $E$ is tightly bound as $ES$).
- $K_m$ (Michaelis Constant): The exact mathematical substrate concentration at which the initial velocity is exactly equal to half of $V_{max}$. It often loosely reflects the enzyme's thermodynamic affinity for the specific substrate (a lower $K_m$ conclusively indicates a much higher affinity).
- $k_{cat}$ (Turnover Number): The total absolute number of individual substrate molecules completely converted to final product per single enzyme molecule per unit time when the enzyme is fully saturated ($V_{max} = k_{cat}[E_{total}]$).
The Lineweaver-Burk Plot (Double Reciprocal)

Figure 6.2: Transformation of the MM equation allows for clear visual identification of inhibition types.
Because estimating the exact asymptote of $V_{max}$ on a hyperbolic curve is difficult, kineticists transform the Michaelis-Menten equation into a linear form by taking the reciprocal of both sides. This results in the Lineweaver-Burk equation:
This equation follows the straight line formula $y = mx + b$, where the y-intercept is $1/V_{max}$ and the x-intercept is $-1/K_m$. This plot makes it exceptionally easy to visually determine the precise type of enzyme inhibition.
6.3 Enzyme Inhibition and Regulation

Kinetics of Enzyme Inhibition: Competitive vs Non-competitive inhibition patterns
Reversible Inhibition
| Inhibition Type | Binding Site | Effect on $V_{max}$ | Effect on $K_m$ | Key Characteristics |
|---|---|---|---|---|
| Competitive | Active Site | No Change | Increases (apparent) | Inhibitor resembles substrate. Can be overcome by massively increasing $[S]$. Lines cross at Y-axis. |
| Uncompetitive | Allosteric Site (only on $ES$) | Decreases | Decreases | Inhibitor binds only after the substrate has bound. Cannot be overcome by $[S]$. Lines are parallel. |
| Mixed | Allosteric Site (on $E$ or $ES$) | Decreases | Changes (usually increases) | Inhibitor binds to a site distinct from the active site. Lines cross to the left of the Y-axis. |
Irreversible Inhibition
Irreversible inhibitors tightly bind to the enzyme, often forming a permanent covalent bond with a crucial amino acid residue in the active site, completely destroying catalytic activity. A highly specialized class is Suicide Inhibitors (Mechanism-based inactivators). These are unreactive compounds that initially bind like normal substrates. However, the normal catalytic mechanism of the enzyme transforms the molecule into a highly reactive intermediate that irreversibly covalently bonds with the enzyme, "killing" it in the process (e.g., Penicillin targeting bacterial transpeptidase).
6.4 Classification of Enzymes
The International Union of Biochemistry and Molecular Biology (IUBMB) classifies all enzymes into six primary classes based on the specific type of chemical reaction they catalyze.
1. Oxidoreductases
Catalyze oxidation-reduction reactions (transfer of electrons or hydride ions). Ex: Dehydrogenases.
2. Transferases
Catalyze the transfer of functional groups (e.g., methyl, phosphate) from one molecule to another. Ex: Kinases.
3. Hydrolases
Catalyze cleavage of bonds by the addition of water (hydrolysis). Ex: Proteases, Lipases.
4. Lyases
Catalyze the cleavage of C-C, C-O, or C-N bonds by means other than hydrolysis or oxidation, often leaving double bonds.
5. Isomerases
Catalyze structural rearrangements within a single molecule (isomerization). Ex: Mutases.
6. Ligases
Catalyze the formation of bonds (C-C, C-S, C-O, C-N) by condensation reactions coupled to ATP cleavage. Ex: DNA Ligase.
6.5 Environmental Effects on Enzyme Activity

Figure 6.3: Optimal ranges for temperature and pH. Deviations lead to structural denaturation or active site ionization changes.
Enzymes are highly sensitive to their physical environment. Both temperature and pH profoundly affect catalytic efficiency:
- Temperature: As temperature increases, the kinetic energy of molecules increases, leading to more frequent collisions between enzyme and substrate, thus increasing reaction rate. However, beyond a specific optimal temperature (often ~37°C in humans), the weak non-covalent bonds stabilizing the enzyme's 3D structure break, causing denaturation and a catastrophic drop in activity.
- pH: Every enzyme has an optimal pH (e.g., Pepsin at pH 2, Trypsin at pH 8). Extreme shifts in pH alter the ionization state of crucial amino acid side chains within the active site, disrupting substrate binding and catalysis, and can also lead to enzyme denaturation.
Regulatory Enzymes: In complex metabolic pathways, specific enzymes can be highly regulated via multiple elaborate mechanisms to control entire pathway flux. These strictly include:
- Allosteric Regulation: Reversible, noncovalent binding of specific regulatory compounds (allosteric modulators) that structurally alter the enzyme's conformation. Allosteric enzymes do NOT follow Michaelis-Menten kinetics (they uniquely show sigmoidal velocity curves).
- Reversible Covalent Modification: Precise addition or complete removal of specific chemical groups (like massive, highly charged phosphate groups added by kinases) that drastically and fundamentally alter entire enzyme activity states.
- Proteolytic Cleavage: Some enzymes are systematically synthesized as entirely inactive precursors (zymogens) and are permanently activated only by highly specific, targeted proteolytic cleavage of precise peptide bonds (e.g., powerful digestive enzymes like chymotrypsin, or the massive blood clotting cascade).
Module 7: Nucleotides & Nucleic Acids

The Molecular Repositories of Biological Information & Chemical Energy
Principle 1: Nucleotides are the most versatile molecules in biochemistry. Beyond their role as the monomeric units of polynucleotides (DNA and RNA), they serve as the universal currency of chemical energy (ATP), essential components of enzyme cofactors (NAD+, FAD), and secondary messengers in signal transduction (cAMP).
7.1 Chemical Properties of Nitrogenous Bases
Purine vs Pyrimidine bases (Adenine, Guanine, Cytosine, Thymine, Uracil)

Watson-Crick base pairing rules (A=T and G≡C) in the DNA double helix
Nitrogenous bases are planar, aromatic, and heterocyclic molecules. They are classified into two categories: Purines (Adenine and Guanine) with a fused five- and six-membered ring system, and Pyrimidines (Cytosine, Thymine, and Uracil) with a single six-membered ring.

Figure 7.1: Molecular structures of the five primary nitrogenous bases.
Aromaticity & UV Absorption
The highly delocalized $\pi$-electron system makes these bases aromatic. This allows them to absorb UV light strongly, with a characteristic absorption maximum near 260 nm. This property is the standard laboratory method for quantifying nucleic acids using the Beer-Lambert Law.
Keto-Enol Tautomerism
Bases exist in equilibrium between tautomeric forms (keto/enol and amino/imino). At physiological pH, the keto and amino forms predominate. This is critically important because improper tautomers can lead to mispairing during DNA replication, potentially causing mutations.
7.2 Nucleotide Conformation: Sugar Puckering & Glycosidic Bonds
The 3D conformation of a nucleotide is determined by two major structural constraints: the Sugar Pucker of the pentose ring and the rotation of the N-glycosidic bond.
Figure 7.2: Sugar pucker geometries ($C_2'$-endo vs $C_3'$-endo) and the Syn/Anti glycosidic bond orientations.
| Structural Feature | Options | Biological Relevance |
|---|---|---|
| Sugar Pucker | $C_2' \text{-endo}$ / $C_3' \text{-endo}$ | B-DNA uses $C_2' \text{-endo}$. A-DNA and RNA use $C_3' \text{-endo}$, which reduces the distance between adjacent phosphates. |
| Glycosidic Bond | Syn vs. Anti | Almost all nucleotides in B-DNA are in the Anti conformation. Syn is observed in the purines of left-handed Z-DNA. |
7.3 Polymorphism of the Double Helix (A, B, and Z DNA)

Structural polymorphism of DNA: Comparison of A, B, and Z forms
The DNA double helix is not a rigid structure. Depending on hydration levels, ionic strength, and nucleotide sequence, it can adopt three distinct forms:
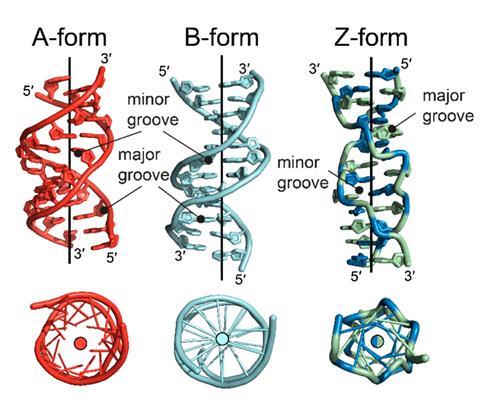
Figure 7.3: Structural comparison of A-form, B-form, and Z-form DNA.
A-Form DNA
- Right-handed
- Short and thick
- $C_3' \text{-endo}$ pucker
- Favored in RNA-DNA hybrids
B-Form DNA
- Right-handed
- Standard biological form
- $C_2' \text{-endo}$ pucker
- Clear Major/Minor grooves
Z-Form DNA
- Left-handed
- Zigzag backbone
- Alternating Anti/Syn
- Favored by GC sequences
7.4 Thermodynamics & The Hyperchromic Effect
The stability of the helix is driven by the Hydrophobic Effect and Base Stacking interactions (Van der Waals/$\pi$-$\pi$ interactions), while hydrogen bonds provide Specificity.

Figure 7.4: DNA melting curve showing the transition to single strands and the increase in UV absorbance.
When DNA is denatured, UV absorbance at 260 nm increases by ~40% (the Hyperchromic Effect) due to the loss of electronic stacking between bases.
The Melting Temperature ($T_m$) is the midpoint of this transition. $T_m$ increases with:
- Higher G-C Content (3 H-bonds vs 2 in A-T, and stronger stacking).
- Higher Ionic Strength (cations mask the repulsion of phosphate backbones).
7.5 DNA Damage & Chemical Mutagenesis
DNA is susceptible to chemical alterations that can result in mutations if not repaired by cellular mechanisms.
Spontaneous Deamination

Deamination involves the loss of an amino group. The most significant biological example is the conversion of Cytosine to Uracil. This is why DNA uses Thymine; it allows repair systems to identify Uracil as a mutation product.
UV-Induced Pyrimidine Dimers

Ultraviolet light induces the formation of covalent Cyclobutane Pyrimidine Dimers (usually between adjacent Thymines). This creates a structural "kink" that stalls replication and transcription.
Polynucleotide Chain Directionality
Module 8: Genes & Chromosomes


Packaging, Topology, and Chromatin Architecture
Principle 1: A biological gene is the fundamental unit of heredity. It is defined as a specific chromosomal DNA segment that encodes a functional product—either a polypeptide or a functional RNA (rRNA, tRNA, snRNA). In eukaryotes, genomic architecture is characterized by a complex mosaic of coding and non-coding sequences.
From Double Helix to Chromosome

Hierarchical Organization: Nucleosomes, solenoids, and higher-order chromatin structure leading to the metaphase chromosome.

Cell Division: Comparative summary of the mechanics and genetic outcomes of Mitosis and Meiosis.
8.1 The Eukaryotic Genome Landscape
Eukaryotic genes are monocistronic but interrupted by non-coding segments. This organization is crucial for the regulation of gene expression and the generation of protein diversity through alternative splicing.

Figure 8.1: Anatomy of a eukaryotic gene, illustrating the relationship between promoters, exons, and introns.
Intron-Exon Organization
- Exons: Coding sequences that are retained in the mature mRNA. Coding regions represent only ~1.5% of the human genome.
- Introns: Intervening sequences removed via splicing. Introns allow for Alternative Splicing, where different combinations of exons are joined to produce distinct proteins.
Non-Coding & Repetitive DNA
The genome is composed largely of repetitive elements:
- Transposons: Mobile DNA elements (~45% of human DNA).
- Regulatory DNA: Distal enhancers and silencers that control transcription.
8.2 DNA Topology and Supercoiling
DNA topology describes the geometric properties of DNA that remain unchanged under continuous deformations. In cellular systems, DNA is typically negatively supercoiled, which facilitates the strand separation required for replication and transcription.

Figure 8.2: The relationship between Linking Number ($L_k$), Twist ($T_w$), and Writhe ($W_r$).
The Fundamental Equation of Topology
$L_k$ is a topological invariant for a closed-circular DNA molecule. Changes in $L_k$ require the breaking and rejoining of phosphodiester bonds, a process mediated by Topoisomerases.
Topoisomerases: Mechanism of Action
Topoisomerases relieve torsional strain (supercoiling) during DNA metabolism. They are classified into two major types based on their catalytic mechanism.

Figure 8.3: Structural mechanism of Type II Topoisomerase, involving a double-strand break and ATP-dependent gate passage.
8.3 Hierarchical Chromatin Organization
Eukaryotic DNA is packaged into chromatin through a hierarchy of structures. The primary unit is the nucleosome, consisting of DNA wrapped around a histone octamer.

Figure 8.4: From the 2 nm DNA helix to the 1400 nm mitotic chromosome—the levels of hierarchical packaging.
- 11 nm Fiber: The "beads-on-a-string" structure. 146 bp of DNA wrapped 1.67 times around a core histone octamer (H2A, H2B, H3, H4).
- 30 nm Fiber: Higher-order coiling stabilized by the linker histone H1.
- Loop Domains: Organized by scaffold proteins (like CTCF) to facilitate gene-specific regulation.
8.4 Heterochromatin vs. Euchromatin
The density of chromatin packaging correlates with gene activity. Transcriptionally active regions are less condensed, while silent regions are highly compacted.

Figure 8.5: Differential staining of Euchromatin (active) and Heterochromatin (silent) within the nucleus.
| Feature | Euchromatin | Heterochromatin |
|---|---|---|
| Packaging State | Decondensed | Condensed |
| Gene Expression | Active | Silent / Repressed |
| Modification State | High Acetylation | High Methylation |
8.5 Specialized Chromosomal Regions
Chromosomes contain specialized regions essential for their stability and segregation during the cell cycle.

Figure 8.6: Detailed anatomy of a chromosome, highlighting centromeres, telomeres, and sister chromatids.
Centromeres
Structural hubs for kinetochore assembly. Centromeres ensure the equal distribution of genetic material by providing the attachment point for spindle fibers during mitosis.
Telomeres
Repetitive DNA caps (TTAGGG) that prevent the loss of genetic information during replication and protect chromosomes from end-to-end fusion.
Epigenetics: The Histone Code
Module 9: DNA Metabolism

Replication, Absolute Fidelity, and the Molecular Machines of Copying
Principle 1: DNA replication is a high-fidelity biological process that ensures the accurate transmission of genetic information. The energetic cost of synthesis is secondary to the massive thermodynamic and enzymatic commitment required to maintain an error rate of less than one per $10^9$ nucleotides.
9.1 The Fundamental Rules of DNA Replication
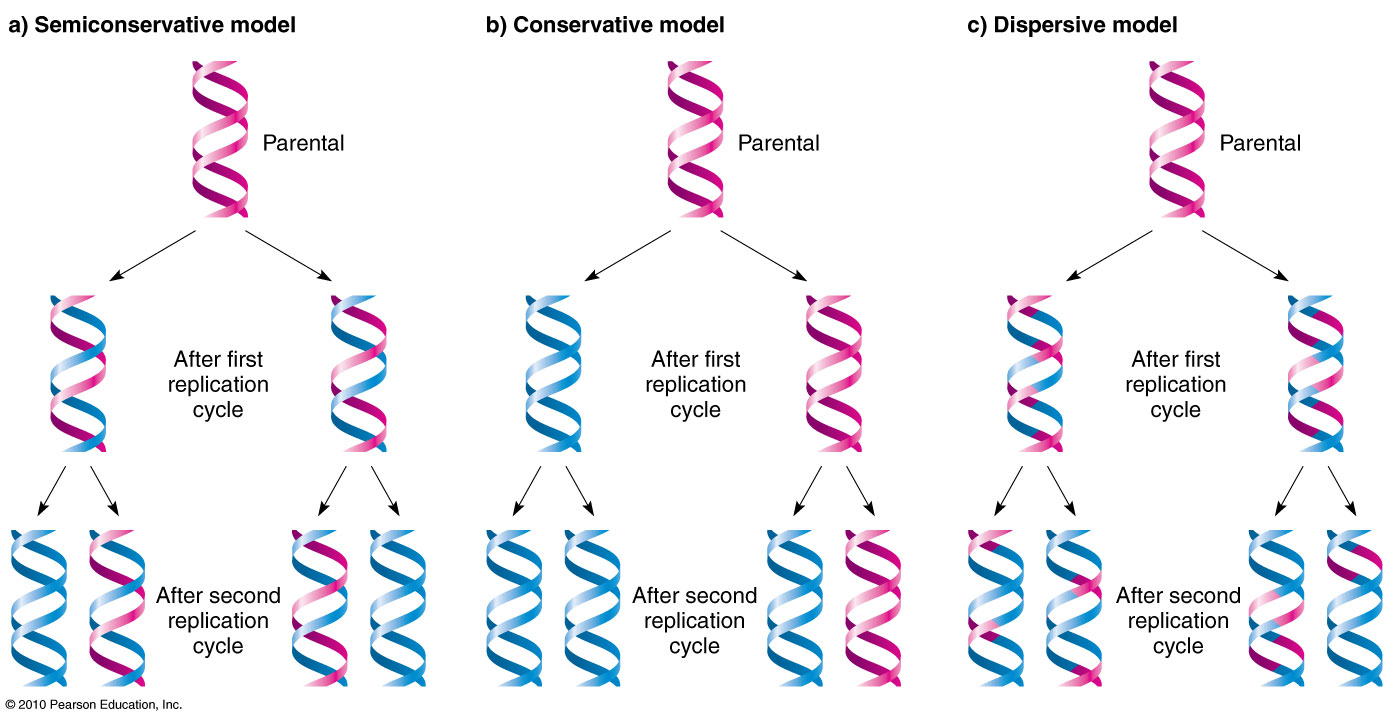
Figure 9.1: The three models of DNA replication. Meselson-Stahl proved the Semiconservative model is correct.
Experimental evidence has established three universal rules for DNA replication:
- Semiconservative: Each daughter helix consists of one parental strand and one newly synthesized daughter strand.
- Bidirectional: Replication starts at unique Origins (ori) and proceeds in both directions simultaneously via two replication forks.
- Semi-discontinuous: Synthesis occurs exclusively 5' → 3'. The Leading Strand is synthesized continuously, while the Lagging Strand is synthesized as Okazaki fragments.
9.2 The Catalytic Mechanism: Two-Metal Ion Catalysis
DNA polymerases do not catalyze the reaction alone; they require two divalent cations (usually $Mg^{2+}$) coordinated by highly conserved Aspartate residues in the active site.

Figure 9.2: Metal ion A facilitates the nucleophilic attack of the 3'-OH, while Metal B stabilizes the pyrophosphate leaving group.
9.3 The Replisome: The Replication Fork Machinery
The Replisome is a massive multi-protein complex that coordinates the unwinding of DNA and the simultaneous synthesis of both strands.

Figure 9.3: Structural orchestration of Helicase, Primase, and Polymerase complexes at the replication fork.
9.4 High Fidelity and Proofreading
Absolute accuracy is achieved through a 3' → 5' Exonuclease activity. When an incorrect base is added, the polymerase stalls and the mismatched end is moved to the exonuclease site for removal.
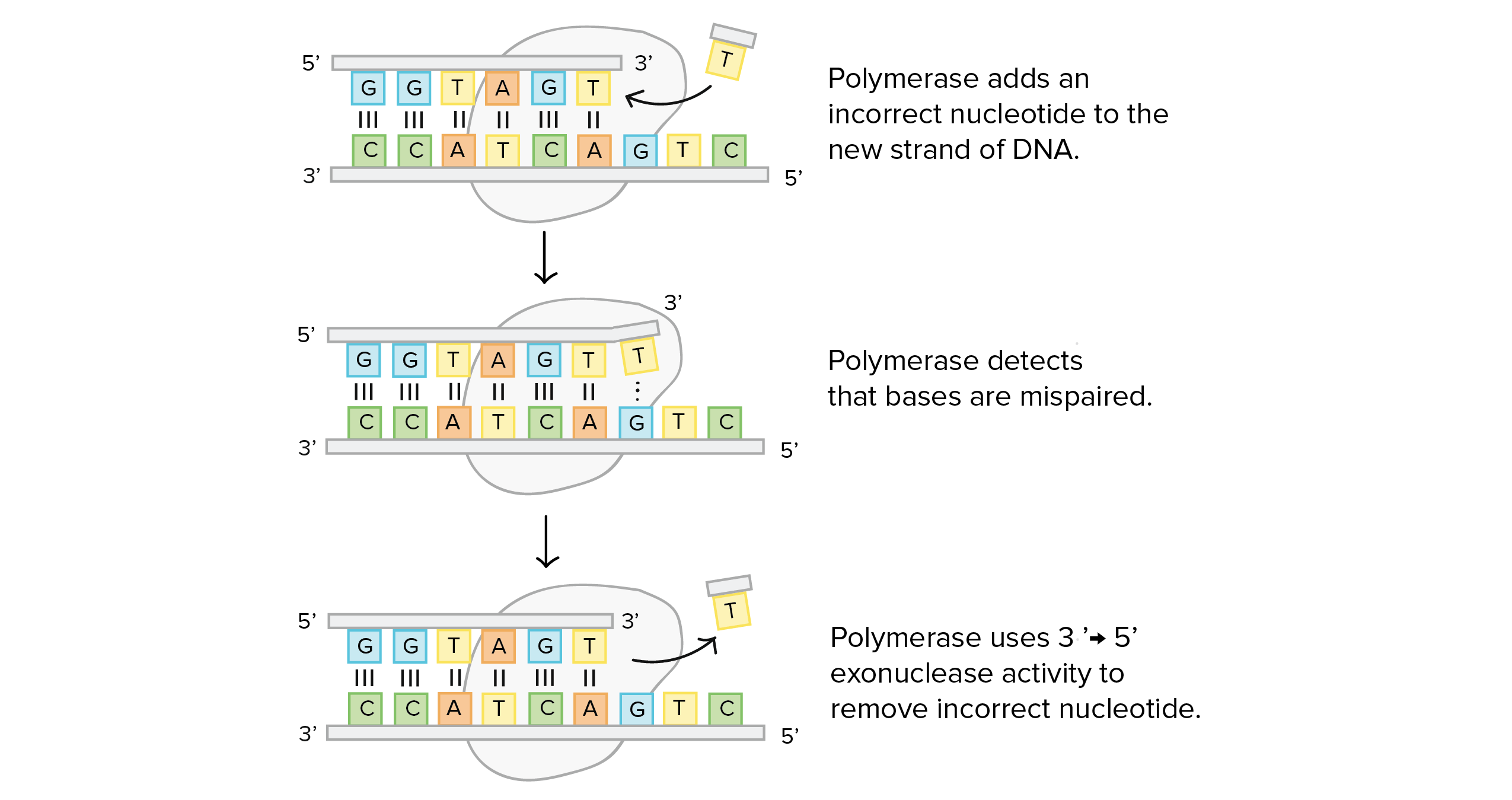
Figure 9.4: The transition between polymerase and exonuclease active sites during proofreading.
9.5 The End-Replication Problem & Telomerase
Linear chromosomes shorten with each round of replication because the lagging strand cannot be completed at the very end. Telomerase solves this by extending the 3' end using its own RNA template.
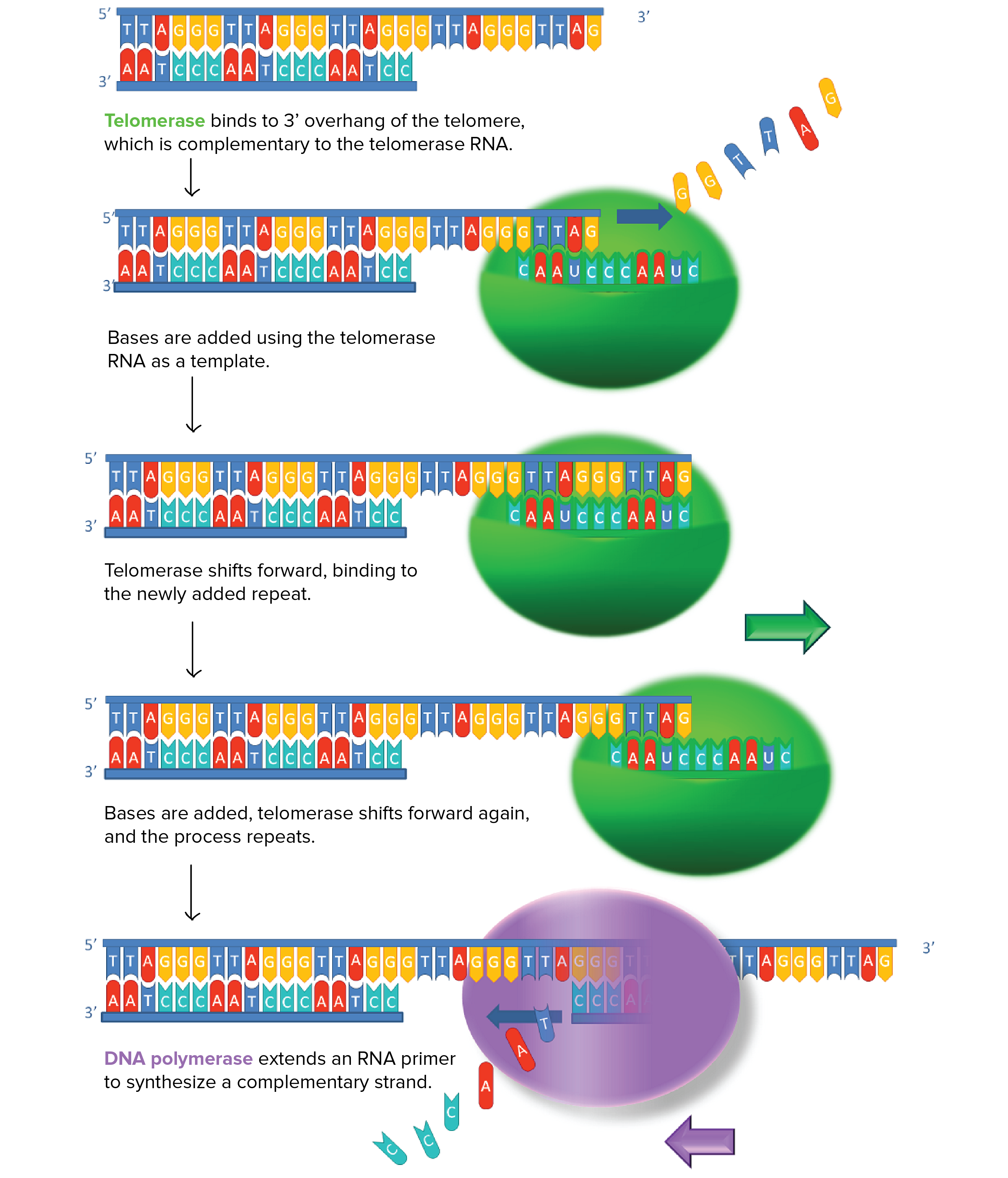
Figure 9.5: Telomerase action prevents the loss of coding DNA by adding repetitive "caps" to chromosome ends.
9.6 DNA Repair Mechanisms
Mismatch Repair (MMR)

MMR systems detect and repair base-pairing errors that bypass proofreading, distinguishing the new strand from the template.
Base Excision Repair (BER)

BER utilizes DNA Glycosylases to excise damaged or unnatural bases, creating an abasic site for repair.
Module 10: RNA Metabolism
Transcription, Massive Processing, and the Precise Flow of Information
Principle 1: Transcription is the first step of gene expression, where a specific segment of DNA is copied into RNA by the enzyme RNA Polymerase. Unlike DNA replication, transcription is selective; only specific genes are transcribed at any given time, a process tightly governed by the interaction between cis-acting elements (promoters/enhancers) and trans-acting factors.
10.1 The Mechanics of Transcription

Figure 10.1: The anatomy of the transcription bubble, showing the DNA-RNA hybrid and polymerase advancement.
The Transcription Process
RNA polymerase unwinds a stretch of DNA (~17 bp) to form a bubble. It catalyzes:
Driven by pyrophosphate hydrolysis, synthesis proceeds 5' → 3' without the need for a primer.
Strand Nomenclature
- Template Strand: Complementary to the nascent RNA.
- Coding (Non-template) Strand: Identical in sequence to the RNA (T replaces U).
10.2 Post-Transcriptional Processing in Eukaryotes
Eukaryotic primary transcripts (hnRNA) must undergo three major modifications within the nucleus to become functional mRNA.
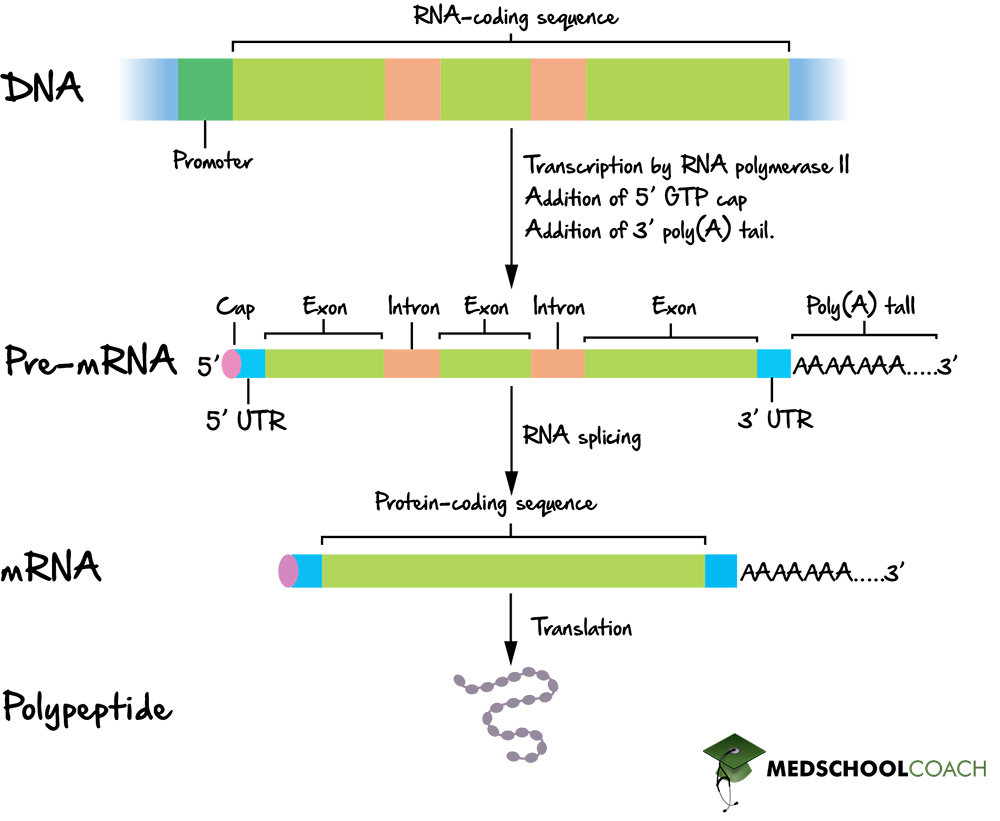
Figure 10.2: The chronological steps of 5' capping, splicing, and 3' polyadenylation.
1. 5' Capping
Addition of a 7-methylguanosine cap via a unique 5'-5' triphosphate linkage to protect against degradation.
2. Splicing
Removal of non-coding introns and ligation of exons, mediated by the Spliceosome (snRNPs).
3. Polyadenylation
Cleavage of the 3' end and addition of a Poly-A tail (~200 As) to enhance mRNA stability and translation.
Module 11: Protein Metabolism
The Universal Genetic Code and Ribosomal Translation
Translation is the conversion of the genetic message into a functional polypeptide. This requires the coordination of mRNA, tRNA, and the enormous Ribosome complex.
11.1 tRNA Activation and the "Second Genetic Code"
The high-fidelity attachment of amino acids to their specific tRNAs is catalyzed by Aminoacyl-tRNA Synthetases (aaRS). This enzymatic accuracy is essential for maintaining the genetic code.

Figure 11.1: The two-step reaction: Amino acid activation (adenylation) followed by tRNA charging.
11.2 The Ribosomal Cycle
The ribosome provides a platform for complementary base pairing between the mRNA codon and the tRNA anticodon, facilitating peptide bond formation.

Figure 11.2: Movement of tRNAs through the Aminoacyl (A), Peptidyl (P), and Exit (E) sites.
- Initiation: Formation of the 70S/80S complex at the start codon (AUG).
- Elongation: tRNA binding, transpeptidation (peptide bond), and translocation.
- Termination: Recognition of stop codons (UAA, UAG, UGA) by Release Factors.
Module 12: Special Topics - Adult Neurogenesis
Hippocampal Plasticity and the Endocannabinoid System
Adult neurogenesis occurs primarily in the Subgranular Zone (SGZ) of the Dentate Gyrus. This dynamic process allows for the continuous integration of new neurons into existing circuits, supporting cognitive flexibility.
12.1 The Neurogenic Lineage
Neural stem cells undergo a defined progression from quiescent precursors to fully mature, integrated granule cells.
Figure 12.1: Developmental stages: Stem cells → Progenitors → Neuroblasts → Mature Neurons.
12.2 Regulation by the Endocannabinoid System (ECS)
The ECS acts as a master rheostat for neurogenesis. Signaling through CB1 and CB2 receptors modulates the proliferation and survival of neural progenitors in response to environmental stimuli.

Figure 12.2: Molecular crosstalk between endocannabinoids and neurogenic pathways.
The "Neurogenesis Hypothesis"
The ECS's ability to drive neurogenesis provides a physiological basis for its effects on mood and learning. Specifically, the anti-depressant effects of exercise are linked to increased endocannabinoid levels, which promote hippocampal cell proliferation.
Module 13: Bonding & Hybridization
The Quantum Mechanical Basis of Organic Structure
Organic chemistry is defined by the unique bonding properties of carbon. Carbon atoms utilize hybridization to achieve optimal spatial orientation for bonding. sp3 hybridization leads to tetrahedral geometry (109.5°), sp2 to trigonal planar (120°), and sp to linear (180°). Single bonds are Sigma (σ) bonds, while double and triple bonds consist of one σ bond and one or two Pi (π) bonds respectively.
Module 14: Isomers & Intermolecular Forces
Molecular Variations and Physical Interactions
Isomers are molecules with the same formula but different structures. Constitutional isomers differ in connectivity, while stereoisomers differ in spatial arrangement. Physical properties like boiling point and solubility are dictated by Intermolecular Forces (IMF), including London dispersion forces, dipole-dipole interactions, and hydrogen bonding.
Module 15: Alkanes & Nomenclature
The Saturated Hydrocarbon Scaffold
Alkanes are saturated hydrocarbons with the general formula $C_nH_{2n+2}$. IUPAC nomenclature provides a systematic way to name complex branched alkanes. The stability of alkane conformations is analyzed using Newman projections, identifying staggered (stable) and eclipsed (unstable) states.
Module 16: Functional Groups
The Reactive Centers of Biomolecules
Functional groups determine the chemical reactivity of organic molecules. Key groups include Hydroxyl (-OH), Carbonyl (C=O), Carboxyl (-COOH), Amino (-NH2), and Phosphate. In biochemistry, the polarity and ionization state of these groups are critical for molecular interactions.
Module 17: Cycloalkanes
Ring Strain and Conformational Analysis
Cycloalkanes experience ring strain due to angle compression and torsional eclipse. Cyclohexane is the most stable, adopting a chair conformation to eliminate strain. Substituents prefer the equatorial position over the axial position to minimize 1,3-diaxial interactions.
Practice Quiz
Test your knowledge on Lehninger Principles