Meditaliano IMAT Prep
Lesson 6: Cell Structure I & Transport
Introduction: The Basic Unit of Life (Advanced)
Welcome to Lesson 6. The cell is the fundamental structural and functional unit of all known organisms. This lesson provides a university-level exploration of the two primary cell architectures—prokaryotic and eukaryotic—before delving into the intricate subsystems that define eukaryotic life: the nucleus, endomembrane system, autonomous organelles, and cytoskeleton.
Learning Objectives (Lesson 6)
- LO 6.1: Differentiate prokaryotic cells (Bacteria, Archaea) from eukaryotic cells, including cell envelope differences (Gram +/-) and the endosymbiotic theory.
- LO 6.2: Detail the structure of the nucleus, including the nuclear pore complex (NPC), Ran-GTP transport cycle, and nuclear lamina.
- LO 6.3: Map the endomembrane system, explaining protein sorting (signal hypothesis), vesicular transport (COPI, COPII, clathrin), and post-translational modifications in the ER and Golgi.
- LO 6.4: Describe the structure and biogenesis of mitochondria and peroxisomes, linking their compartments to their metabolic roles (e.g., cristae for ETC, matrix for Krebs cycle).
- LO 6.5: Compare the structure, dynamics (treadmilling, dynamic instability), and functions of the three major cytoskeleton filaments, including their associated motor proteins (kinesin, dynein, myosin).
- LO 6.6: Differentiate the major types of cell junctions (tight, adherens, desmosomes, gap) and the unique features of plant cells (wall, plasmodesmata, vacuole).
Part 1: The Two Great Cellular Architectures
The primary division of life is between Prokaryotes (Bacteria and Archaea) and Eukaryotes (Eukarya). This division is based on fundamental differences in cellular organization, most notably the eukaryotic cell's compartmentalization through a true nucleus and membrane-bound organelles.
1.1 The Prokaryotic Cell
Prokaryotic cells are structurally simpler but metabolically diverse. They lack a nucleus and extensive internal membranes.
- Nucleoid: The region (not an organelle) where the single, circular chromosome is located, highly supercoiled and organized by nucleoid-associated proteins (NAPs).
- Cell Envelope: The structures enclosing the cytoplasm. In bacteria, this is key for classification:
- Gram-positive: Thick peptidoglycan cell wall external to a single plasma membrane.
- Gram-negative: A thin peptidoglycan wall located in the periplasmic space between the inner and outer plasma membranes.
- Cytoskeleton: Prokaryotes possess simple cytoskeletal homologs, such as FtsZ (a tubulin homolog for cell division) and MreB (an actin homolog for cell shape).
Image 1: The Two Great Cellular Architectures

Visual Analysis: The Two Great Cellular Architectures
Based on LO 6.1, this visual compares prokaryotic and eukaryotic architectures and the origin of eukaryotes through endosymbiosis.
- Prokaryotic vs. Eukaryotic Comparison:
- Prokaryotes (Left): Focus on the nucleoid (circular DNA), FtsZ/MreB cytoskeleton homologs, binary fission, and bacterial cell wall details (Gram-positive thick peptidoglycan vs. Gram-negative double membrane and periplasm).
- Eukaryotes (Right): True nucleus (histones, linear chromosomes), complex cytoskeleton (microtubules, IFs, actin), complex endomembrane systems, and differences between animal/plant cells.
- Endosymbiotic Theory (Bottom Right): Pathways for the engulfment of α-proteobacteria (mitochondria) and cyanobacteria (chloroplasts). Key evidence includes double membranes, circular DNA, 70S ribosomes, and binary fission.
1.2 The Eukaryotic Cell & Endosymbiotic Theory
Eukaryotic cells are defined by their complex compartmentalization. This complexity is believed to have arisen, in part, from endosymbiosis, a theory proposing that mitochondria and chloroplasts were once free-living prokaryotes engulfed by an ancestral host cell.
Diagram of an Animal Cell

Diagram of a Plant Cell

Evidence for Endosymbiotic Theory
- Mitochondria and chloroplasts have their own circular DNA, similar to prokaryotes.
- They possess 70S ribosomes, smaller than the 80S ribosomes in the eukaryotic cytoplasm and identical in size to prokaryotic ribosomes.
- They are enclosed by a double membrane (the inner membrane being the original prokaryote's, the outer from the host's vesicle).
- They replicate by binary fission, independent of the cell's nuclear division (mitosis/meiosis).
| Feature | Prokaryotes | Eukaryotes |
|---|---|---|
| Nucleus | Absent. Genetic material is in a nucleoid region. | Present. A double membrane encloses the DNA. |
| Membrane-Bound Organelles | Absent (no mitochondria, ER, Golgi, etc.). | Present (mitochondria, ER, Golgi, lysosomes, etc.). |
| DNA Structure | Single, circular chromosome. | Multiple, linear chromosomes complexed with histone proteins. |
| Size | Typically small (1-5 μm). | Typically larger (10-100 μm). |
| Ribosomes | Smaller (70S). | Larger (80S in cytoplasm; 70S in mitochondria/chloroplasts). |
| Cytoskeleton | Simple (FtsZ, MreB). | Complex (Actin, Intermediate Filaments, Microtubules). |
| Cell Division | Binary Fission. | Mitosis and Meiosis. |
Part 2: The Information Core: Nucleus & Ribosomes
2.1 The Nucleus: Structure and Transport
The nucleus is the defining feature of the eukaryotic cell. It protects the genome and isolates the processes of transcription and RNA processing from translation.
- Nuclear Envelope: A double membrane (outer and inner) separated by a perinuclear space. The outer membrane is continuous with the endoplasmic reticulum.
- Nuclear Lamina: A meshwork of lamin proteins (a type of intermediate filament) on the inner surface of the nuclear envelope. It provides structural support and helps organize chromatin.
- Nuclear Pore Complex (NPC): An intricate, massive protein channel (made of nucleoporins) that regulates all traffic into and out of the nucleus. Small molecules diffuse freely, but large molecules (like proteins and RNA) require active, selective transport.
Image 2: The Nucleus and Intracellular Traffic

Visual Analysis: The Nucleus and Intracellular Traffic
This image details the information core of the cell and its transport mechanisms (LO 6.2, 6.3).
- Nuclear Structure: High-resolution look at the Nuclear Pore Complex (NPC), the double-membrane envelope (Outer/Inner/Perinuclear), and the structural support provided by the Nuclear Lamina (Lamin proteins).
- The Ran-GTP Cycle (Bottom Left): A step-by-step visualization of protein import (NLS-protein + Importin), cargo release via Ran-GTP binding, and the recycling of transport receptors. The spatial roles of Ran-GEF (nuclear) and Ran-GAP (cytoplasmic) are highlighted.
- Endomembrane Flow (Right): Comprehensive mapping of vesicular traffic between the RER, SER, and Golgi. Key features include COPII (anterograde), COPI (retrograde), and Clathrin (lysosomal/endocytic) coats, as well as protein modifications like N-linked (ER) and O-linked (Golgi) glycosylation.
Nuclear Transport: The Ran-GTP Cycle
Protein import into the nucleus is a key process. A protein with a Nuclear Localization Signal (NLS) binds to a transport receptor called importin. This complex moves through the NPC. Inside the nucleus, a small G-protein called Ran-GTP binds to importin, causing it to release the cargo protein. The importin-Ran-GTP complex is then exported back to the cytoplasm, where GTP is hydrolyzed to GDP, releasing importin for another cycle.
2.2 The Nucleolus and Ribosome Biogenesis
The nucleolus is a non-membranous structure within the nucleus, visible as a dense spot. It is the site of ribosome biogenesis.
- It consists of three regions: the Fibrillar Center (FC), Dense Fibrillar Component (DFC) where rRNA is transcribed, and the Granular Component (GC) where ribosomal subunits are assembled.
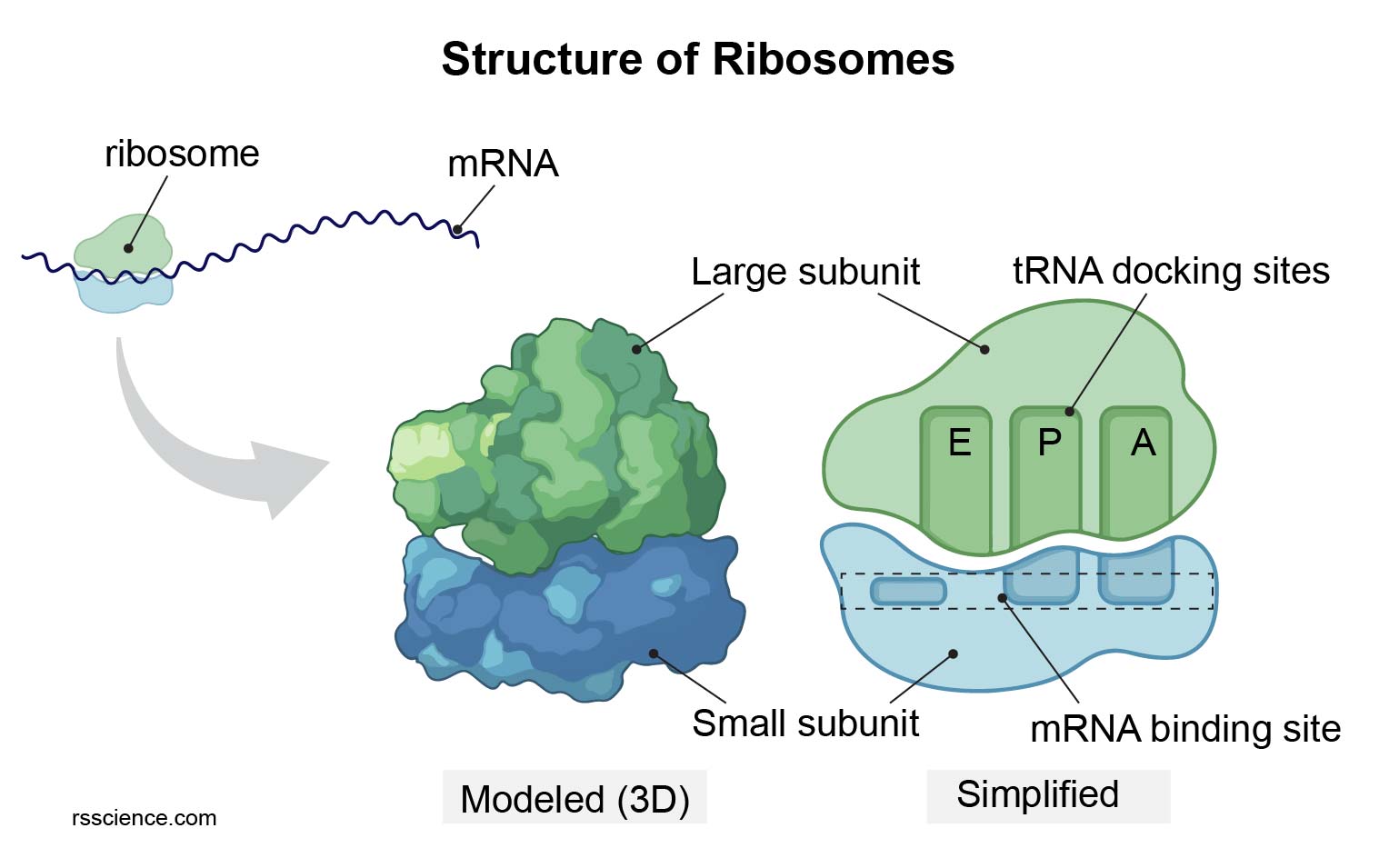
- Eukaryotic ribosomes are 80S, composed of a 60S large subunit and a 40S small subunit. They are assembled in the nucleolus and exported to the cytoplasm.
- Ribosomes translate mRNA into protein, operating at three sites: the A site (aminoacyl-tRNA), P site (peptidyl-tRNA), and E site (exit).
Structure of a Ribosome
Part 3: The Endomembrane System: A Cellular Factory
This system is a dynamic, interconnected network of membranes responsible for protein synthesis, modification, packaging, and transport, as well as lipid synthesis.
3.1 Endoplasmic Reticulum (ER)
The ER is a vast network of cisternae and tubules continuous with the outer nuclear membrane.
- Rough ER (RER): Studded with ribosomes. As proteins are synthesized (co-translational import), they enter the ER lumen, where they are folded by chaperone proteins (like BiP), undergo N-linked glycosylation, and are subject to quality control. Misfolded proteins trigger the Unfolded Protein Response (UPR).
- Smooth ER (SER): Lacks ribosomes. Its functions include: 1) Lipid synthesis (phospholipids, steroids), 2) Detoxification of drugs and poisons (e.g., via cytochrome P450 enzymes), and 3) Calcium sequestration (storage), which is critical for cell signaling (e.g., sarcoplasmic reticulum in muscle).
Image 6: Protein Sorting & ER Quality Control

Visual Analysis: Protein Sorting & ER Quality Control
This image provides a sophisticated look at the "Quality Control" systems of the Endoplasmic Reticulum (LO 6.3).
- Co-translational Processing: Details on N-linked glycosylation and the chaperones (BiP, Calnexin, Calreticulin) that guide nascent polypeptide chains into their functional three-dimensional shapes.
- ERAD Pathway: The mechanism for ER-Associated Degradation. Misfolded proteins are "retrotranslocated" back to the cytoplasm, tagged with ubiquitin by E3 ligases, and destroyed by the proteasome.
- UPR Signaling (Bottom): The three branches of the Unfolded Protein Response (ATF6, IRE1, PERK) are visualized. These sensors monitor ER stress and coordinate a response that either restores homeostasis (by increasing chaperone production) or triggers apoptosis if the damage is irreparable.
3.2 The Golgi Apparatus: Sorting and Shipping
The Golgi consists of a stack of flattened, membrane-bound sacs called cisternae. It functions as a "post office," receiving cargo from the ER and modifying, sorting, and packaging it for its final destination. This occurs via the cisternae maturation model, where cisternae themselves move and mature from the *cis* (entry) face to the *trans* (exit) face.
- Cis Golgi Network (CGN): Receives vesicles from the ER.
- Medial & Trans Cisternae: Sites of further protein modification, such as O-linked glycosylation and modification of N-linked glycans.
- Trans Golgi Network (TGN): The final sorting station. Proteins are packaged into different vesicles for delivery to the plasma membrane (secretion), lysosomes, or other organelles.
The Endomembrane System (ER, Golgi, Vesicles)

3.3 Vesicular Transport and Lysosomes
Transport between these compartments is mediated by vesicles, which are "tagged" by specific coat proteins:
- COPII-coated vesicles: Anterograde transport (ER to Golgi).
- COPI-coated vesicles: Retrograde transport (Golgi back to ER, or within Golgi).
- Clathrin-coated vesicles: Transport from the TGN to endosomes/lysosomes, and in receptor-mediated endocytosis.
Vesicle fusion with the target membrane is mediated by SNARE proteins (v-SNAREs on the vesicle, t-SNAREs on the target membrane).
Lysosomes are the cell's digestive compartments, containing acid hydrolases. They are formed when transport vesicles from the TGN (tagged with a mannose-6-phosphate signal) fuse with late endosomes.
Part 4: Energy & Metabolism Hubs
4.1 Mitochondria
The mitochondrion is the primary site of cellular respiration and ATP synthesis. It is a dynamic organelle that undergoes constant fusion and fission.
- Outer Membrane: Contains porins, large channel proteins that allow free passage of small molecules.
- Intermembrane Space: The region between the membranes. Protons ($H^+$) are pumped here by the ETC. Contains cytochrome c, a key factor in initiating apoptosis (programmed cell death).
- Inner Membrane: Highly folded into cristae, dramatically increasing surface area. It is rich in the unique phospholipid cardiolipin and is impermeable to ions. It houses the Electron Transport Chain (ETC) and ATP Synthase.
- Matrix: The innermost compartment. Contains enzymes for the Krebs Cycle and fatty acid $\beta$-oxidation, as well as mitochondrial DNA (mtDNA) and 70S ribosomes.
Image 3: Energy & Metabolism Hubs

Visual Analysis: Energy & Metabolism Hubs
Based on LO 6.4, this visual summarizes the structure and specialized roles of the cell's metabolic powerhouses.
- Mitochondria (Left): A detailed look at the double-membrane system. Highlighted features include the inner membrane's cristae (housing the ETC and ATP Synthase), the unique lipid cardiolipin, and the matrix containing the Krebs cycle enzymes, 70S ribosomes, and circular mtDNA. Dynamics of fusion and fission are also represented.
- Peroxisomes (Right): Focus on the single-membrane structure and the critical role of Catalase in neutralizing $H_2O_2$. Key reactions include the $\beta$-oxidation of very-long-chain fatty acids (VLCFA) and detoxification of substances like alcohol. Biogenesis from the ER or existing peroxisomes is shown.
4.2 Chloroplasts
Found in plants and algae, chloroplasts are the site of photosynthesis. They are a type of plastid.
- Three-Membrane System: An outer membrane, inner membrane, and the internal thylakoid membrane system.
- Thylakoids: Flattened, interconnected sacs where the Light-Dependent Reactions occur. Stacks of thylakoids are called grana.
- Stroma: The fluid-filled space outside the thylakoids, analogous to the mitochondrial matrix. It contains enzymes for the Calvin Cycle, chloroplast DNA, and 70S ribosomes.
Structure of a Chloroplast

4.3 Peroxisomes
Peroxisomes are small, single-membrane metabolic compartments that are involved in diverse oxidative reactions. They acquire proteins from the cytosol and can arise from ER budding or by fission of existing peroxisomes.
- Beta-oxidation: They break down very-long-chain fatty acids.
- Detoxification: They neutralize toxic substances (like alcohol) by transferring hydrogen to oxygen, forming hydrogen peroxide ($H_2O_2$).
- Catalase: They contain the enzyme catalase, which immediately breaks down the toxic $H_2O_2$ into water and oxygen.
Structure of a Peroxisome

Part 5: The Cytoskeleton: Internal Scaffolding
The cytoskeleton is a dynamic network of protein fibers responsible for cell shape, internal organization, and movement.
| Filament | Monomer | Structure | Dynamics & Polarity | Key Functions |
|---|---|---|---|---|
| Microfilaments | G-Actin | Two intertwined strands (F-Actin) | Polar (+ and - ends). Exhibits treadmilling. | Cell cortex, muscle contraction (with myosin), microvilli, cell motility (lamellipodia), cytokinesis (contractile ring). |
| Microtubules | $\alpha/\beta$-Tubulin Dimer | Hollow tube of 13 protofilaments | Polar (+ and - ends). Exhibits dynamic instability (GTP cap). | Organelle transport (tracks for motors), mitotic spindle, core of cilia and flagella (9+2 axoneme). Organized by MTOCs. |
| Intermediate Filaments | Various (e.g., Keratin, Vimentin, Lamins) | Rope-like fibers (coiled-coils) | Non-polar. Less dynamic. | Mechanical strength, bearing tension, anchoring nucleus (lamina), cell-cell junctions (desmosomes). |
Image 4: The Dynamic Cytoskeleton & Motors

Visual Analysis: The Dynamic Cytoskeleton & Motors
Based on LO 6.5, this image details the internal scaffolding of the cell and the engines that power cellular movement.
- Filament Architectures (Top):
- Actin (Microfilaments): Visualizing G-actin monomers assembling into the F-actin double helix and the phenomenon of treadmilling powered by ATP hydrolysis.
- Intermediate Filaments: Representation of rope-like, non-polar fibers that provide immense mechanical strength to the cell.
- Microtubules: Details on the $\alpha/\beta$-tubulin dimers forming a 13-protofilament tube, showing dynamic instability driven by the GTP cap.
- Motor Dynamics (Bottom):
- Muscle Contraction: A high-resolution sliding filament model showing the interaction between actin and myosin.
- Intracellular Transport: Mapping kinesin movement toward the plus-end and dynein toward the minus-end of microtubule tracks.
- Ciliary Motion: A cross-section of the 9+2 axoneme structure, detailing how dynein arms and nexin links coordinate the bending of cilia and flagella.
Motor Proteins: The Engines of the Cell
Motor proteins use energy from ATP hydrolysis to move along cytoskeletal "tracks".
- Myosins: Move along actin filaments (e.g., in muscle contraction and cytokinesis).
- Kinesins: Most move toward the plus-end of microtubules (anterograde transport, e.g., moving vesicles to the cell periphery).
- Dyneins: Move toward the minus-end of microtubules (retrograde transport, e.g., moving vesicles to the cell center). Also responsible for the bending of cilia and flagella.
Motor Proteins (Kinesin and Dynein) on a Microtubule

Part 6: Cell Junctions, Walls, and ECM (Advanced)
6.1 Animal Cell Junctions
In tissues, cells are connected to each other and to the extracellular matrix (ECM) by specialized junctions.
- Tight Junctions (Zonula Occludens): Formed by claudins and occludins. They seal the space between epithelial cells, preventing paracellular transport (leakage between cells).
- Adherens Junctions (Zonula Adherens): Formed by cadherins. They link the actin cytoskeletons of adjacent cells, forming an "adhesion belt".
- Desmosomes (Macula Adherens): Formed by cadherins (desmoglein, desmocollin). They link the intermediate filaments of adjacent cells, providing strong, spot-like adhesion.
- Gap Junctions: Formed by connexin proteins. They create channels that allow for the direct passage of small molecules and ions between adjacent cells, coupling them metabolically and electrically.
Types of Animal Cell Junctions

6.2 Plant Cell Structures
Plant cells have unique structures reflecting their sessile, autotrophic lifestyle.
- Cell Wall: A rigid outer layer composed of cellulose microfibrils embedded in a matrix of hemicellulose and pectin. Provides structural support and protection.
- Central Vacuole: A very large organelle (up to 90% of cell volume) enclosed by a membrane called the tonoplast. It stores water, maintains turgor pressure, and often contains digestive enzymes (acting as a lysosome).
- Plasmodesmata: Channels that pass through the cell walls of adjacent plant cells, connecting their cytoplasm. They are the functional equivalent of gap junctions.
Part 7: The Plasma Membrane and Transport
7.1 The Fluid Mosaic Model Revisited
The fluid mosaic model describes the plasma membrane as a fluid structure with a "mosaic" of various proteins embedded in or attached to a bilayer of phospholipids. The membrane is selectively permeable. Cholesterol molecules help maintain membrane fluidity at different temperatures.
Image 5: Advanced Cell Structures & Transport

Visual Analysis: Advanced Cell Structures & Transport
This integrated visual brings together the complex structural and functional features of both animal and plant cells (LO 6.6, 7).
- Animal Cell Junctions (Top): Comprehensive mapping of the four major junctions. Tight junctions are shown sealing the paracellular space, while Adherens and Desmosomes provide mechanical linkage to the cytoskeleton. Gap junctions are visualized as channels for metabolic coupling.
- Plant Cell Architecture (Bottom Left): Detailed structure of the rigid cell wall (cellulose/pectin), the pressure-regulating central vacuole, and the plasmodesmata that allow for cytoplasmic continuity.
- Integrated Transport (Bottom Right): A master overview of membrane transport. Includes passive mechanisms (simple/facilitated diffusion, osmosis) and active systems (Primary $Na^+/K^+$ pump, Secondary symport). Bulk transport via exocytosis and the various forms of endocytosis (phagocytosis/pinocytosis) are also detailed.
7.2 Passive Transport: No Energy Required
Passive transport is the diffusion of a substance across a membrane with no energy investment. Substances move down their concentration gradient.
- Simple Diffusion: The net movement of small, nonpolar molecules like $O_2$, $CO_2$, and lipids directly through the lipid bilayer.
- Facilitated Diffusion: The passage of polar molecules (like glucose) and ions (like $Na^+$) with the help of specific transmembrane transport proteins (channels or carriers).
- Osmosis: The diffusion of free water across a selectively permeable membrane.
Osmosis in Animal Cells (RBCs)

In hypotonic solution, animal cells lyse (burst). In hypertonic, they shrivel.
Osmosis in Plant Cells

Plants prefer hypotonic (turgid). In hypertonic, they undergo plasmolysis.
7.3 Active Transport: Energy Required
Active transport uses energy (usually ATP) to move solutes against their concentration gradient.
• Primary Active Transport: Directly uses ATP. The sodium-potassium pump is a key example (3 $Na^+$ out, 2 $K^+$ in).
• Secondary Active Transport (Cotransport): Uses the concentration gradient created by a primary active transporter.
Diagram: Sodium-Potassium Pump (Primary Active Transport)

7.4 Bulk Transport: Vesicles for Large Molecules
Large molecules cross the membrane in bulk via vesicles, which requires energy.
- Exocytosis: The cell secretes molecules by the fusion of vesicles with the plasma membrane.
- Endocytosis: The cell takes in molecules by forming new vesicles. Includes phagocytosis ("cellular eating"), pinocytosis ("cellular drinking"), and receptor-mediated endocytosis (highly specific).
Bulk Transport: Endocytosis and Exocytosis

Part 8: Acellular Structures - Viruses (Advanced)
Viruses are infectious particles that lie on the border of life. They are obligate intracellular parasites, meaning they lack the machinery for self-replication and must hijack a host cell's resources to reproduce. They are not considered cells and thus fall outside the cell theory.
Why aren't viruses alive?
Because they have no cellular organelles, no independent metabolism, and cannot reproduce on their own outside a host. The simplicity and parasitic nature of viruses differentiate them fundamentally from living cells.
Viral Structure
- Genetic Material: The viral genome can be DNA or RNA, single- or double-stranded.
- Capsid: A protein shell that encloses and protects the genome.
- Envelope (Optional): Many viruses have a lipid envelope derived from the host cell membrane, which contains viral glycoproteins used for attachment.
Basic Structure of a Virus

Interactive Practice Quiz (Lesson 6)
Test your understanding of all topics (Advanced Structure, Junctions, Transport, Viruses). Choose the best answer for each question and then submit to see your results.