Meditaliano Lesson 9 : Bioenergetics
🔥 0. Thermodynamics & Bioenergetics Fundamentals
Bioenergetics is the rigorous quantitative study of energy transductions within living cells. It is governed strictly by the fundamental Laws of Thermodynamics. Understanding these underlying physical principles is an absolute prerequisite, as they are frequently tested in the IMAT integrated science section to evaluate a student's grasp of interdisciplinary concepts linking biology and physics.
The Universal Laws of Thermodynamics
-
First Law (Conservation of Energy): Energy can neither be created nor destroyed out of nothing. It can only change forms.
Biological Application: In biological systems, this means the massive chemical potential energy stored in the covalent bonds of a glucose molecule is entirely accounted for: a fraction of it is successfully transformed into the chemical energy of ATP phosphoanhydride bonds, while the inevitable remainder is dissipated irrevocably into the environment as thermal energy (heat).
-
Second Law (Entropy): In every physical or chemical change occurring spontaneously, the universe always tends toward greater disorder or randomness. This measure of disorder is known as Entropy ($S$).
Biological Implication: Cells are highly ordered, highly complex, low-entropy structures. To maintain this local order against the relentless laws of physics, a cell must constantly undergo highly exergonic catabolic reactions (like cellular respiration). These reactions release massive amounts of heat, thereby increasing the entropy of the surrounding universe enough to satisfy the Second Law overall. Without this constant energy expenditure, the cell succumbs to entropy and dies.
Gibbs Free Energy ($\Delta G$)
Gibbs Free Energy is defined as the amount of energy available to do useful biological work at a constant temperature and pressure. The defining thermodynamic equation is essential for the IMAT:
Where $\Delta H$ is the change in enthalpy (the total heat content of the system), $T$ is the absolute temperature in Kelvin, and $\Delta S$ is the change in entropy. Based strictly on the value of $\Delta G$, biochemical reactions are classified into two critical categories for metabolism:
| Reaction Type | $\Delta G$ Sign | Spontaneity & Energy Flow | Biological Examples |
|---|---|---|---|
| Exergonic | < 0 (Negative) | Spontaneous (Releases Free Energy) | Cellular Respiration, ATP Hydrolysis, Catabolic breakdown of macromolecules. |
| Endergonic | > 0 (Positive) | Non-spontaneous (Requires Energy Input) | Photosynthesis, Protein Synthesis, DNA Replication, Active Transport, Anabolism. |
| Equilibrium | = 0 (Zero) | No net work can be done by the system | A biological cell at true thermodynamic equilibrium is a dead cell. |
This is the fundamental principle of all cellular metabolism. Cells cannot simply "push" an endergonic (energy-requiring) reaction forward against the laws of physics. Instead, they brilliantly pair a highly exergonic reaction (almost always the hydrolysis of ATP) with the required endergonic reaction (like building a complex peptide bond). As long as the overall combined sum of $\Delta G$ for the coupled system remains mathematically negative, the entire combined process proceeds spontaneously.
🔋 1. ATP: The Universal Energy Currency
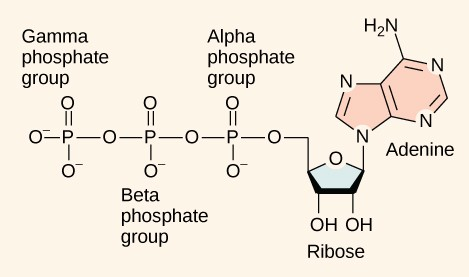
Adenosine Triphosphate (ATP) is structurally an RNA nucleoside triphosphate. It is the primary energy-transfer molecule in all domains of life on Earth, acting as a ubiquitous, rapidly rechargeable battery that powers almost every endergonic cellular process.
Structural Components of the ATP Molecule:
- Adenine: A complex, double-ringed purine nitrogenous base.
- Ribose: A 5-carbon pentose sugar. (Note: The specific combination of Adenine bonded to Ribose is termed Adenosine).
- The Triphosphate Tail: Three distinct phosphate groups attached in series to the 5' carbon of the ribose sugar ring. They are designated as $\alpha$ (closest to the ribose), $\beta$ (the middle phosphate), and $\gamma$ (the terminal phosphate). The high-energy covalent bonds connecting these specific phosphate groups are referred to as phosphoanhydride bonds.
Why are Phosphoanhydride bonds considered "High Energy"?
The phrase "high energy bond" is frequently misunderstood; it is a historical misnomer. The energy does not reside physically within the electron orbitals of the bond itself. Instead, the "high energy" refers exclusively to the massive difference in Gibbs free energy between the highly unstable reactant state and the highly stable product state.
At standard physiological pH (around 7.4), the triphosphate tail of ATP is fully ionized, carrying four closely packed, dense negative charges. These localized negative charges violently repel each other, creating immense electrostatic repulsion. It is mechanistically identical to trying to hold a tightly compressed, heavy-duty mechanical spring. Cleaving the terminal $\gamma$-phosphate via hydrolysis relieves this massive structural tension. Furthermore, the resulting products (ADP and inorganic phosphate, $P_i$) undergo immediate resonance stabilization and highly favorable hydration by surrounding water molecules. This makes the product state significantly lower in potential energy than the reactant state, thus releasing a massive, sudden burst of free energy capable of doing work.
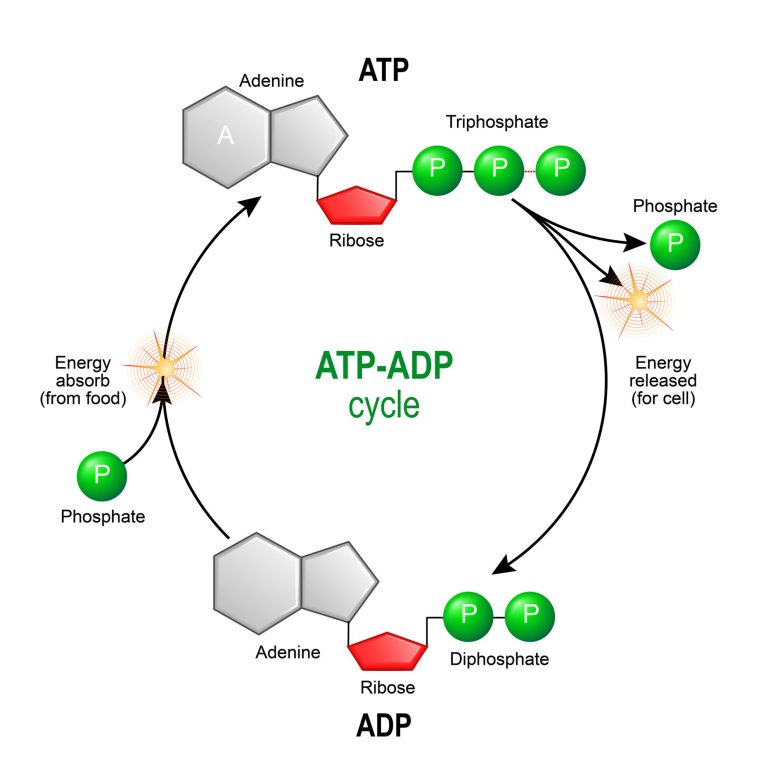
The ATP-ADP Cycle and Exergonic Hydrolysis
When the terminal $\gamma$-phosphate bond is broken by the enzymatic addition of a water molecule (hydrolysis), ATP is converted into ADP (Adenosine Diphosphate) and a free inorganic phosphate ion ($P_i$). This specific hydrolysis reaction provides the requisite driving energy for cellular mechanical, transport, and chemical work.
$\Delta G^{\circ'} = -7.3 \text{ kcal/mol} \text{ (or } -30.5 \text{ kJ/mol)}$
Because the cellular supply of ATP is extremely limited, the cell must continuously and rapidly recycle ADP back into ATP. It achieves this by utilizing the energy harvested from the slow, oxidative breakdown of food molecules during cellular respiration. This continuous loop is the ATP-ADP Cycle. The turnover rate in human biology is staggering; a single, actively contracting muscle cell can recycle its entire internal pool of ATP molecules in less than one minute.
⚙️ 2. Enzyme Kinetics and Mechanisms
Every metabolic pathway, including every single sequential step of cellular respiration and photosynthesis, is rigidly governed by enzymes. Enzymes are biological catalysts, overwhelmingly composed of globular proteins (with the exception of some specialized RNA ribozymes), that vastly accelerate the rate of chemical reactions without being permanently altered or consumed in the process.
Mechanism of Action: Lowering the Activation Energy
Enzymes speed up reactions strictly and exclusively by lowering the Activation Energy ($E_a$) required for the reactants to reach the transition state. The transition state is the highly unstable, peak-energy configuration of the reactant molecules. By binding the substrates and physically stabilizing this transition state, enzymes allow the reaction to proceed millions or even billions of times faster. Crucially for the IMAT: Enzymes DO NOT change the overall $\Delta G$ (free energy change) of the reaction, nor do they ever alter the final equilibrium constant ($K_{eq}$).
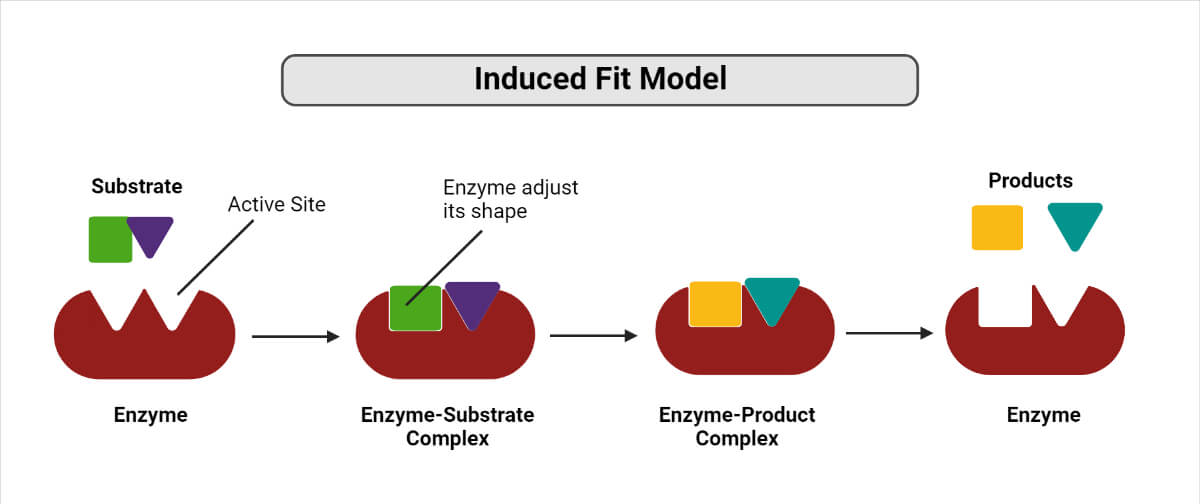
The Modern Induced-Fit Model
The outdated historical "Lock and Key" model incorrectly assumed the enzyme's active site was perfectly rigid and pre-formed. Modern biochemistry relies instead on the Induced-Fit Model. In this dynamic model, the active site is inherently flexible. When the specific substrate enters the active site, the enzyme undergoes a distinct conformational (shape) change to mold tightly and perfectly around the substrate. This tight structural embrace applies physical, mechanical, and chemical stress to specific covalent bonds within the substrate, forcibly bending them towards the transition state and dramatically facilitating the reaction.
Cofactors and Coenzymes: The Essential Helpers
Many complex enzymes are synthesized by the ribosome as completely inactive proteins, referred to as Apoenzymes. They absolutely require the binding of non-protein helper molecules to achieve their proper 3D conformation and become fully functional, catalytically active Holoenzymes.
- Cofactors: These are typically inorganic metallic ions that bind tightly to the enzyme to assist directly in electron transfer or to provide crucial structural stabilization to the active site. Common IMAT examples include Magnesium ($Mg^{2+}$) strictly required by Hexokinase and all other kinases, Iron ($Fe^{2+}$) in cytochromes, and Zinc ($Zn^{2+}$) in DNA polymerase.
- Coenzymes: These are complex organic helper molecules. In human biology, they are vastly overwhelmingly derived from vitamins consumed in the diet. They act as transient, mobile carriers of specific functional groups or high-energy electrons between different enzymes.
- $NAD^+$ is derived from Vitamin B3 (Niacin).
- FAD is derived from Vitamin B2 (Riboflavin).
- Coenzyme A (CoA) is derived from Vitamin B5 (Pantothenic acid).
⚡ 3. Cellular Respiration: The Big Picture
Cellular respiration is the master catabolic, exergonic pathway of the eukaryotic cell. It involves the highly controlled, stepwise oxidation of organic monomers (such as glucose, fatty acids, and amino acids) to extract their usable chemical energy. If a cell were to simply burn glucose in a single, uncontrolled explosive step (like a fire), the activation energy would be insurmountable without high heat, and once overcome, all the energy would be released instantly as blinding light and destructive heat, instantly incinerating the cell. Instead, cells utilize a multi-step enzymatic cascade to lower activation energies and harvest the energy slowly and safely in small packets, trapping it efficiently in the form of ATP.
The Central Role of Electron Carriers in Redox Reactions
Cellular respiration is fundamentally a long series of redox (reduction-oxidation) reactions. Oxidation involves the loss of electrons, while reduction is the gain of electrons (Remember the essential mnemonic OIL RIG: Oxidation Is Loss, Reduction Is Gain). In biological systems, tracking electrons often simply means tracking Hydrogen atoms, because a standard hydrogen atom consists of exactly one proton ($H^+$) and one electron ($e^-$).
- Throughout the various stages of respiration, the Glucose molecule is heavily and repeatedly oxidized, eventually losing all of its hydrogen atoms and electrons to become low-energy $CO_2$ gas.
- Oxygen gas ($O_2$), possessing the highest electronegativity of any biological molecule, serves as the final, ultimate destination. It is completely reduced by gaining those electrons and protons to become highly stable water ($H_2O$).
Instead of transferring high-energy electrons directly from glucose to oxygen—which would be explosive—the cell employs intermediate "electron shuttles" or "taxis". These are the soluble dinucleotide coenzymes:
It accepts exactly 2 electrons and 1 proton to become the high-energy reduced form NADH.
It accepts exactly 2 electrons and 2 protons to become the high-energy reduced form FADH$_2$.
These reduced coenzymes ($NADH$ and $FADH_2$) physically carry the harvested high-energy electrons from the cytoplasm and the mitochondrial matrix directly to the Electron Transport Chain embedded on the inner mitochondrial membrane.
The Three Main Stages and Mitochondrial Geography
Aerobic respiration is highly compartmentalized to maximize efficiency and maintain concentration gradients. It is universally divided into four distinct stages:
Image 1: Complete Overview of Cellular Respiration

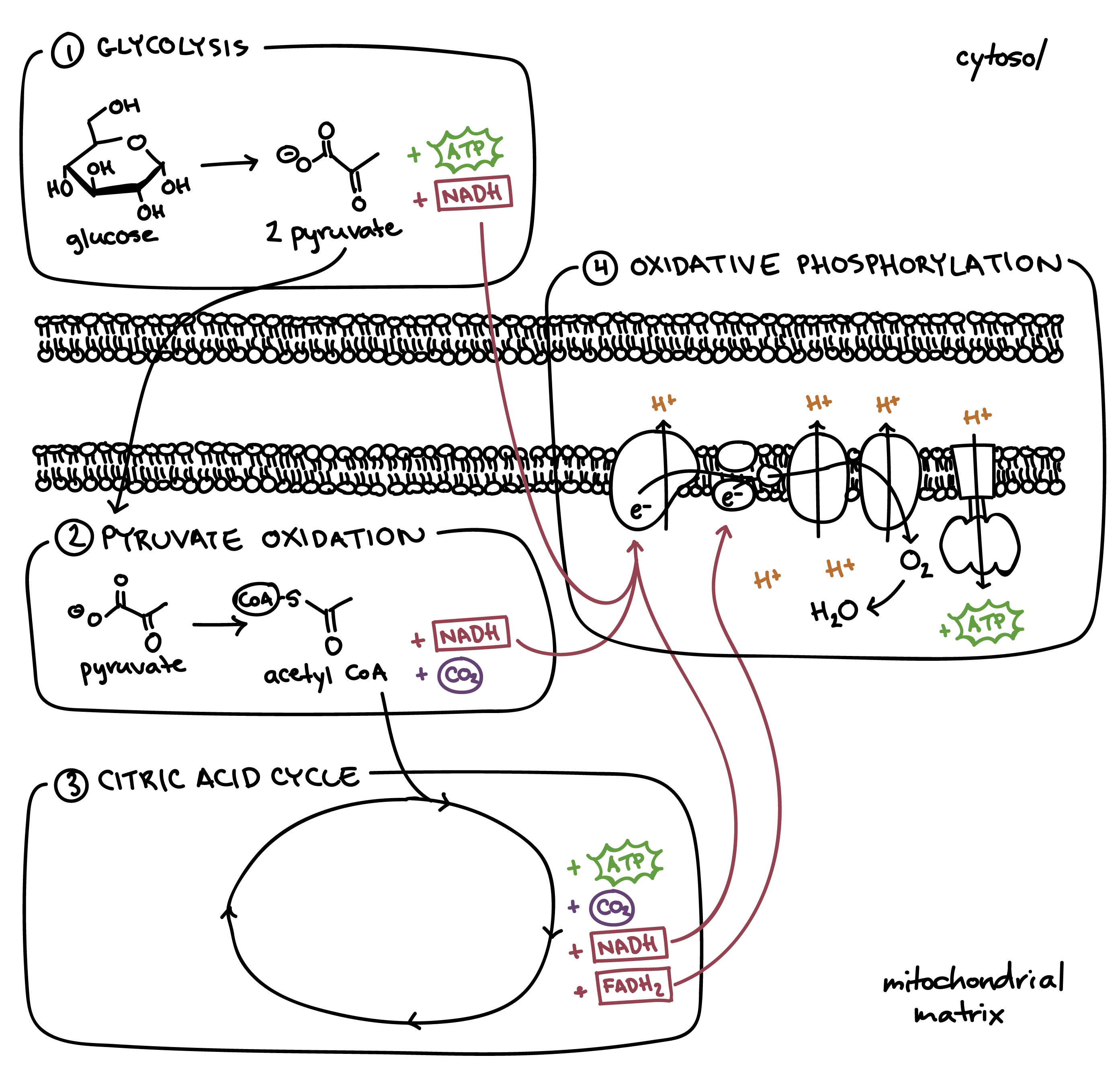
Integrated Metabolic Flow
This diagram tracks the standard carbon skeleton (6C → 3C → 2C → waste) and all energy yields (ATP/NADH/FADH$_2$) through the system:
- Compartmentalization: Clearly distinguishes between cytosolic events (Glycolysis) and mitochondrial matrix/membrane events.
- Carbon Tracking: Illustrates the complete breakdown to 6 $CO_2$ molecules.
- Energy Totals: Culminates in the generation of a theoretical maximum of 32 ATP per glucose molecule.
To fully grasp the later, high-yield stages of respiration, an IMAT candidate must thoroughly understand the specific structure of the mitochondrion. It is a double-membraned organelle specifically evolved for the sole purpose of advanced bioenergetics.


2. Inner Membrane: Highly impermeable and strictly selective. It contains the massive, embedded protein complexes of the Electron Transport Chain and ATP Synthase.
3. Cristae: Deep, intricate folds of the inner membrane. These folds exponentially increase the surface area available for oxidative phosphorylation, allowing tens of thousands of ETC copies to exist per mitochondrion.
4. Matrix: The dense, viscous internal fluid containing the enzymes for Pyruvate Oxidation, the entirety of the Krebs Cycle, and the Beta-Oxidation of fatty acids. It also houses the circular mitochondrial DNA and 70S ribosomes.
5. Intermembrane Space: The narrow, confined gap between the inner and outer membranes. This is the crucial reservoir where protons ($H^+$) are pumped and concentrated by the ETC, creating a highly acidic, low-pH environment essential for driving chemiosmotic ATP synthesis.
🔢 4. Glycolysis ("Sweet Splitting")
Glycolysis is an ancient, highly conserved 10-step metabolic pathway that enzymatically cleaves one 6-carbon glucose molecule into two 3-carbon pyruvate molecules. Because it does not require oxygen, it serves as the foundational pathway for both aerobic respiration and anaerobic fermentation. The pathway is conceptually divided into two distinct halves: an Energy Investment Phase (Steps 1-5, which consumes ATP to prime and destabilize the sugar) and an Energy Payoff Phase (Steps 6-10, which yields a net profit of ATP and NADH).
Phase 1: Preparatory / Energy Investment Phase
In this initial phase, the cell spends 2 ATP molecules to sequentially phosphorylate the glucose molecule, raising its free energy state and making the ring structure unstable enough to be cleaved in half.
-
Hexokinase (or Glucokinase in liver cells).
Phosphorylation: Glucose entering the cell via GLUT transporters is immediately phosphorylated on its 6th carbon to become Glucose-6-Phosphate (G6P).
Consumes 1 ATP.Biological purpose: The addition of the bulky, highly negatively charged phosphate group traps the glucose inside the cell, as the charged molecule can no longer fit back through the hydrophobic interior of the cell membrane transport proteins. This is a strictly irreversible step requiring $Mg^{2+}$ as a necessary cofactor to shield the negative charges of ATP. -
Phosphoglucose Isomerase
Isomerization: The six-membered pyranose ring of G6P (which is an aldohexose) is enzymatically rearranged into a five-membered furanose ring, creating Fructose-6-Phosphate (F6P, a ketohexose). This critical rearrangement moves the carbonyl oxygen, preparing the molecule for symmetric cleavage. -
Phosphofructokinase-1 (PFK-1)
The Committed Step: A second phosphate group is forcefully added to carbon 1, forming the highly unstable, nearly symmetrical Fructose-1,6-bisphosphate (F1,6BP).
Consumes 1 ATP. (Total spent: 2 ATP).High Yield IMAT Concept: Regulation PFK-1 is universally considered the most important rate-limiting enzyme and the primary thermodynamic control point of the entire glycolytic pathway. It is subject to strict, complex allosteric regulation to match the cell's energy demands.- Inhibited by: High intracellular levels of ATP (indicating energy abundance) and high levels of Citrate (indicating that the Krebs cycle in the mitochondria is backed up and does not need more fuel).
- Activated by: High levels of AMP (indicating severe energy depletion).
-
Aldolase
Lysis (Cleavage): The highly unstable 6-carbon F1,6BP is enzymatically cleaved down the middle into two distinct, but interconvertible, 3-carbon isomers: Dihydroxyacetone phosphate (DHAP) and Glyceraldehyde-3-phosphate (G3P). -
Triose Phosphate Isomerase (TIM)
Isomerization: The downstream enzymes of glycolysis are highly specific and can only recognize and process G3P. Therefore, TIM rapidly and reversibly converts the DHAP into a second molecule of G3P. The reaction is continuously pulled forward towards G3P according to Le Chatelier's principle because G3P is immediately consumed by the next step.
Crucial Milestone: From this exact point forward, every single chemical reaction occurs TWICE for every one original molecule of glucose.
Phase 2: Energy Payoff Phase
Reminder: All steps listed below happen exactly $\times 2$ per original glucose molecule!
-
Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH)
Oxidation & Phosphorylation: G3P is oxidized. The extracted high-energy electrons (and a proton) are transferred to $NAD^+$, reducing it to form NADH. Simultaneously, the massive energy released from this exergonic oxidation is used to forcibly attach a free inorganic phosphate ($P_i$) floating in the cytosol onto the molecule, forming the high-energy intermediate 1,3-Bisphosphoglycerate (1,3-BPG).
Yields 1 NADH (x2 = 2 NADH total).Note: This is the ONLY redox (oxidation-reduction) reaction in the entire glycolysis pathway. Crucially, note that it utilizes free inorganic phosphate ($P_i$), not ATP, to add the second phosphate group. -
Phosphoglycerate Kinase
First Substrate-Level Phosphorylation: The newly added, extremely high-energy phosphate on carbon 1 of 1,3-BPG is transferred directly to an ADP molecule, forging a new phosphoanhydride bond to form ATP. The remaining molecule is now 3-Phosphoglycerate (3PG).
Yields 1 ATP (x2 = 2 ATP total). Metabolic Accounting: This specific step achieves the thermodynamic "breakeven" point, exactly repaying the 2 ATP debt incurred during Phase 1. -
Phosphoglycerate Mutase
Phosphate Shift: The remaining phosphate group is enzymatically relocated from Carbon 3 to Carbon 2, forming 2-Phosphoglycerate (2PG). This shift destabilizes the molecule, preparing it for the upcoming dehydration reaction. -
Enolase
Dehydration: The enzyme removes a molecule of water ($H_2O$) from 2PG. This removal creates a carbon-carbon double bond, resulting in the formation of Phosphoenolpyruvate (PEP).
Biochemical significance: This simple dehydration vastly redistributes energy within the molecule, creating a massive high-energy enol-phosphate bond. PEP is an incredibly potent phosphate donor, possessing a higher phosphoryl-transfer potential than ATP itself. (Clinical Note: Fluoride is a well-known competitive inhibitor of enolase, which is why fluoride is added to blood sample tubes to stop glycolysis and preserve blood glucose levels for testing). -
Pyruvate Kinase (PK)
Second Substrate-Level Phosphorylation: PEP transfers its high-energy enol-phosphate directly to ADP, forming another molecule of ATP and yielding the final 3-carbon stable product of the pathway, Pyruvate.
Yields 1 ATP (x2 = 2 ATP total). This is the final, highly exergonic, essentially irreversible regulatory step of glycolysis. The ATP generated in this step represents the actual net profit of ATP for the cell.
Image 2: Glycolysis ("Sweet Splitting") - A Deep Dive

The 10-Step Enzymatic Cascade
This deep-dive visual separates the pathway into two distinct functional phases:
- Energy Investment (Steps 1–5): Details the PFK-1 "Committed Step" and the initial expenditure of 2 ATP molecules.
- Energy Payoff (Steps 6–10): Tracks the production of 4 ATP and 2 NADH molecules. Note that this phase occurs twice per original glucose.
- Key Features: Includes all 10 enzyme names (e.g., Hexokinase, Enolase) and highlights the critical molecular split at G3P.
Glycolysis Net Mass Balance & Energy Accounting
| Input (per 1 Glucose molecule) | Output (per 1 Glucose molecule) | Net Profit for the Cell |
|---|---|---|
| 1 Glucose (6C) | 2 Pyruvate (3C) | 2 Pyruvate |
| 2 ATP (invested) | 4 ATP (gross) | + 2 ATP |
| 2 NAD⁺ | 2 NADH + 2 H⁺ | + 2 NADH |
🔗 5. Pyruvate Oxidation (The Link Reaction)
Under aerobic conditions, the pyruvate generated in the cytosol cannot be processed further there. It must be actively transported across the double membrane of the mitochondria via a specific $H^+$/pyruvate symporter. Once inside the highly controlled environment of the Mitochondrial Matrix, it encounters one of the largest, most structurally intricate multi-enzyme complexes in eukaryotic biology: the Pyruvate Dehydrogenase Complex (PDC). The PDC serves as the crucial biochemical bridge connecting glycolysis to the Krebs cycle.
The 3 Catalytic Steps of the PDC Reaction:
The PDC consists of three distinct enzymes (E1, E2, E3) working in tight spatial coordination to execute the following sequence:
- 1. Decarboxylation (E1 enzyme): The 3-carbon pyruvate molecule is stripped of its carboxyl group. This group is released as a molecule of $CO_2$ gas. This represents a major metabolic milestone: the very first time that carbon originating from the original glucose molecule is fully oxidized and released as metabolic waste.
- 2. Oxidation (E2 enzyme): The remaining 2-carbon fragment (a hydroxyethyl group) is severely oxidized into an acetyl group. The high-energy electrons extracted during this oxidation are passed through an internal relay of covalently bound cofactors (first to lipoic acid, then to FAD) and finally deposited onto a mobile $NAD^+$, forming a molecule of high-energy NADH.
- 3. CoA Attachment (E2 enzyme): The newly formed, highly reactive acetyl group is covalently attached to a large carrier molecule called Coenzyme A (CoA) via a highly unstable, high-energy sulfur bond (thioester bond), forming the final, transportable product: Acetyl-CoA (2C).
PDC Cofactors & Severe Neurological Diseases
The Pyruvate Dehydrogenase Complex is extraordinarily large and complex. To function properly, it absolutely requires the presence of 5 specific vitamin-derived coenzymes. A genetic or dietary deficiency in any one of these cripples the entire complex. The medical mnemonic to remember them for the IMAT is "TLC For Nancy":
- Thiamine Pyrophosphate (TPP / derived from Vitamin B1)
- Lipoic Acid (synthesized internally, but critical for E2)
- CoA (derived from Pantothenic Acid / Vitamin B5)
- FAD (derived from Riboflavin / Vitamin B2)
- NAD$^+$ (derived from Niacin / Vitamin B3)
A severe dietary deficiency in Thiamine (Vitamin B1) leads to debilitating diseases like Beriberi or Wernicke-Korsakoff syndrome (which is extremely common in cases of chronic alcoholism). Without a functioning PDC due to lack of TPP, pyruvate backs up in the cytosol and cannot enter the Krebs cycle. It is instead forcibly shunted into the lactic acid fermentation pathway, causing severe, life-threatening lactic acidosis. Furthermore, because brain tissue relies almost exclusively on the aerobic oxidation of glucose for energy, PDC failure leads to rapid neurological damage, confusion, and eventually collapse.
Image 3: Pyruvate Oxidation (The Link Reaction) & The 5 Cofactors

Molecular Bridge to the Matrix
This advanced card details the exactly 3-step mechanism inside the mitochondrial matrix:
- Step-by-Step Flow: 1. Decarboxylation (loss of $CO_2$), 2. Oxidation (reduction of $NAD^+$), and 3. CoA Attachment (thioester bond formation).
- The 5 Essential Cofactors: Summarizes the "TLC For Nancy" mnemonic (TPP, Lipoic Acid, CoA, FAD, $NAD^+$) and their vitamin precursors.
- Clinical Integration: Includes a dedicated box linking PDC failure to Wernicke-Korsakoff syndrome and Beriberi, highlighting the neurological impact of B1 deficiency.
(Calculated per original Glucose molecule = 2 Pyruvates processed)
🔄 6. The Krebs Cycle (TCA / Citric Acid Cycle)
The Krebs cycle constitutes the central metabolic hub of the cell. It completes the exhaustive breakdown of glucose molecules that was initiated in glycolysis by fully oxidizing the 2-carbon acetyl group of Acetyl-CoA into two molecules of low-energy $CO_2$. It is defined formally as a closed "cycle" or "loop" because the final enzymatic step perfectly regenerates the exact 4-carbon compound (Oxaloacetate) that is required to condense with Acetyl-CoA to begin the first step.
Crucially for stoichiometric accounting: Because one glucose molecule yields two Acetyl-CoA molecules via the link reaction, the Krebs cycle must turn exactly TWICE for every original molecule of glucose.
The 8 Intermediates Mnemonic
"Can I Keep Selling Sex For Money, Officer?"
Image 4: The Krebs Cycle (TCA / Citric Acid Cycle) - A Complete Enzymatic Map

Carbon Oxidation and Energy Yields
This comprehensive map is designed for rote memorization and tracks every high-energy yield point:
- Step-by-Step Substrates: Details the 6C → 5C → 4C carbon loop and the regeneration of oxaloacetate.
- Energy Yield points: Clearly marks where 3 NADH, 1 FADH$_2$, 1 direct ATP (via GTP), and 2 $CO_2$ are generated per single turn.
- Regulatory Callouts: Highlights strict allosteric regulation and the cycle's "Amphibolic" nature, where intermediates feed into other synthetic pathways.
The 8 Enzymatic Steps in Deep Detail
-
Citrate Synthase
Condensation: The 2-carbon acetyl group from Acetyl-CoA is transferred to the 4-carbon acceptor molecule, Oxaloacetate (OAA). This condensation reaction forms a 6-carbon molecule, Citrate (hence the alternative name, Citric Acid Cycle). The massive Coenzyme A molecule is released intact, returning to the link reaction to pick up another acetyl group. This step is highly exergonic and virtually irreversible, pulling the cycle forward. -
Aconitase
Isomerization: The tertiary alcohol group of citrate is structurally poorly suited for the upcoming oxidative decarboxylations. Aconitase rearranges the molecule by removing a water molecule (forming cis-aconitate) and then immediately adding it back in a different stereochemical position, converting Citrate into its isomer, Isocitrate (6C).
Toxicology Note: Fluoroacetate (a highly toxic pesticide and rat poison) is converted by citrate synthase into fluorocitrate, which acts as a deadly suicide inhibitor of aconitase, shutting down the entire Krebs cycle and killing the organism. -
Isocitrate Dehydrogenase
Oxidative Decarboxylation 1: Major Rate-Limiting Step. Isocitrate (6C) undergoes severe oxidation, transferring high-energy electrons to $NAD^+$ to yield the cycle's first NADH. The resulting unstable intermediate immediately loses a carboxyl group as $CO_2$ gas. The molecule has now been degraded to a 5-carbon compound: $\alpha$-Ketoglutarate. -
$\alpha$-Ketoglutarate Dehydrogenase Complex
Oxidative Decarboxylation 2: This massive enzyme complex is structurally and functionally homologous to the PDC (requiring the exact same 5 "TLC For Nancy" cofactors). $\alpha$-Ketoglutarate (5C) loses another carbon as $CO_2$, reduces a second $NAD^+$ to NADH, and attaches a new Coenzyme A via a high-energy thioester bond to form Succinyl-CoA (4C).
Metabolic Milestone: At this exact point, two carbon atoms have entered the cycle from Acetyl-CoA, and two carbon atoms have left the cycle as $CO_2$. The glucose molecule has now been completely and fully oxidized. The remaining four steps of the cycle serve solely to rearrange the remaining 4-carbon skeleton back into Oxaloacetate to keep the cycle spinning. -
Succinyl-CoA Synthetase
Substrate-Level Phosphorylation: The high-energy thioester bond connecting CoA to the succinyl group is enzymatically cleaved. The tremendous energy released is harnessed to directly phosphorylate a GDP molecule into GTP. (In cells, GTP is energetically equivalent to ATP and is rapidly converted into ATP by nucleoside diphosphate kinase). The remaining 4-carbon skeleton is Succinate. -
Succinate Dehydrogenase
Oxidation: Succinate (4C) is oxidized into Fumarate (4C) by the creation of a double bond. Because the change in free energy is too small to reduce $NAD^+$, the extracted electrons are instead transferred to a tightly bound FAD molecule, creating FADH$_2$.
IMAT Alert - Crucial Structural Exception:Unlike all other Krebs cycle enzymes which float freely in the aqueous matrix, Succinate Dehydrogenase is an integral membrane protein tightly embedded deep within the inner mitochondrial membrane. It serves double duty: it is the step 6 enzyme of the Krebs cycle AND it is simultaneously Complex II of the Electron Transport Chain, physically feeding its FADH$_2$ electrons directly into the lipid bilayer.
-
Fumarase
Hydration: A molecule of water is added across the trans double bond of Fumarate, stereospecifically creating L-Malate (4C). -
Malate Dehydrogenase
Final Oxidation: Malate (4C) is oxidized back into Oxaloacetate (4C). The extracted electrons reduce a final $NAD^+$ into NADH. The cycle is now closed, and the regenerated Oxaloacetate is ready to condense with a new Acetyl-CoA molecule.
Krebs Cycle Net Yield & Mass Balance Accounting
| Metabolic Product | Yield per 1 Acetyl-CoA (1 Turn of Cycle) |
Total Yield per 1 Glucose (2 Turns of Cycle) |
|---|---|---|
| NADH | 3 (Steps 3, 4, 8) |
6 |
| FADH₂ | 1 (Step 6) |
2 |
| ATP (via GTP) | 1 (Step 5) |
2 |
| CO₂ Gas | 2 (Steps 3, 4) |
4 (Exhaled as waste) |
⚡ 7. Electron Transport Chain & Oxidative Phosphorylation
This final, magnificent stage of aerobic respiration is responsible for producing the vast majority (~90%) of the ATP generated from glucose. It is the culmination of all the oxidative work done in the previous stages. Oxidative phosphorylation represents the brilliant biochemical coupling of two distinct, highly coordinated processes: The Electron Transport Chain (which builds a physical, electrochemical battery across the membrane) and Chemiosmosis (which uses the energy of that battery to power a mechanical molecular motor that synthesizes ATP).
Image 5: Electron Transport Chain & Oxidative Phosphorylation: Chemiosmotic Coupling (32 ATP Max)

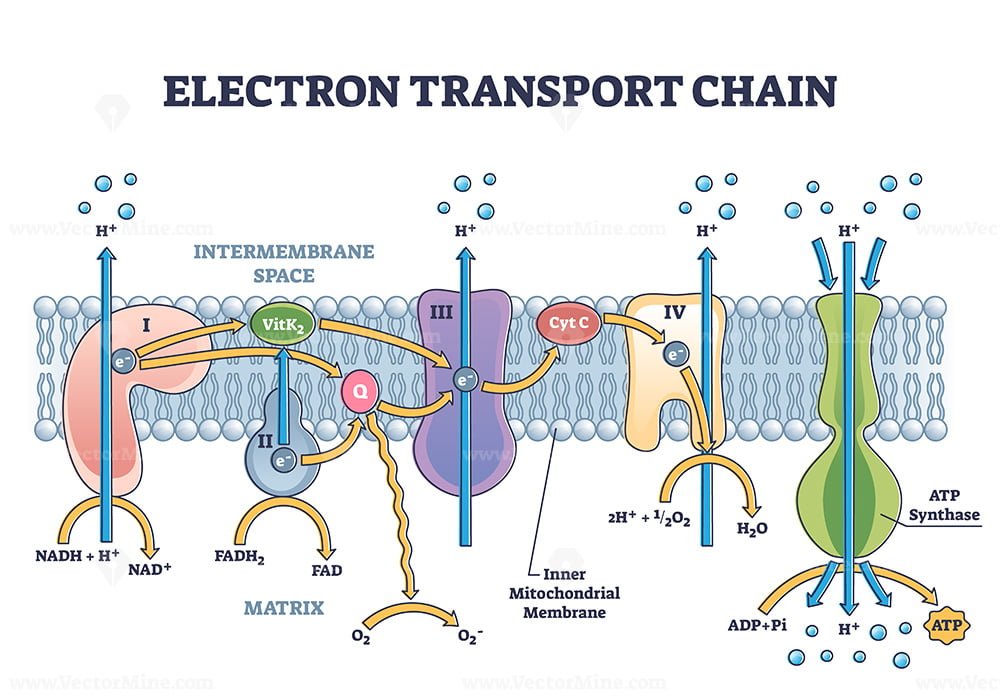
Coupling Oxidation to Phosphorylation
This final comprehensive diagram illustrates the physical battery that drives the cell:
- Energetic Flow: High-energy electrons from 10 NADH and 2 FADH$_2$ flow "downhill" through Complexes I, II, III, and IV to reach Oxygen.
- The Battery (PMF): Highlights the active pumping of protons (4, 4, 2 $H^+$) into the intermembrane space, building the electrochemical gradient.
- Molecular Motor: Details ATP Synthase (Complex V) with its F0 rotor and F1 catalytic knob, showing the mechanical Slamming of ADP and $P_i$.
- Final Yield Accounting: Provides the ultimate ATP calculation based on modern P/O ratios (~2.5 and ~1.5) for a maximum net profit of 32 ATP per glucose.
Detailed Chemiosmotic Coupling & Transmembrane Protein Complexes
A. In-Depth Analysis: The Electron Transport Chain Protein Complexes
The massive protein components of the ETC are arranged strictly and sequentially in order of increasing electronegativity. NADH is a strong electron donor (possessing very low electronegativity and highly negative reduction potential), while Oxygen ($O_2$) is the ultimate, incredibly powerful electron acceptor (highest electronegativity). As electrons "fall" down this energetic staircase, they release energy in discrete, manageable amounts, which is harvested to pump protons.
| Complex Name & Identity | Prosthetic Groups (IMAT Focus) | Biochemical Function | Protons Pumped |
|---|---|---|---|
| Complex I NADH Dehydrogenase |
Contains Flavin Mononucleotide (FMN) and multiple Iron-Sulfur (Fe-S) clusters. | Oxidizes NADH in the matrix. Extracts 2 high-energy electrons and passes them through the Fe-S clusters to Ubiquinone (Q). The massive energy released during this drop is used to pump 4 protons. | 4 H⁺ |
| Complex II Succinate Dehydrogenase |
Contains FAD and Iron-Sulfur (Fe-S) clusters. | The exact same enzyme from Step 6 of the Krebs Cycle. It oxidizes FADH₂. Crucially, it does NOT span the membrane entirely, so it lacks the physical channel to pump protons. It only feeds electrons into Ubiquinone (Q). | 0 H⁺ |
| Ubiquinone (Q) Coenzyme Q10 |
A small, hydrophobic benzoquinone. (It is a lipid, NOT a protein). | A highly mobile, lipid-soluble carrier. It rapidly diffuses entirely within the hydrophobic core of the lipid bilayer, physically carrying electrons from Complexes I & II over to Complex III. | - |
| Complex III Cytochrome bc1 Complex |
Contains Cytochromes b, c1 (heme groups containing Iron), and Rieske Fe-S clusters. | Oxidizes Ubiquinone and reduces Cytochrome c. It utilizes a highly complex, non-linear mechanism called the "Q-cycle" to pump protons efficiently. | 4 H⁺ |
| Cytochrome c Cyt c |
Contains Heme c (an Iron-containing porphyrin ring). | A small, water-soluble peripheral membrane protein. It loosely associates with the outer face (IMS side) of the inner membrane, shuttling electrons one at a time from Complex III to Complex IV. | - |
| Complex IV Cytochrome c Oxidase |
Contains Cytochromes a, a3, and critical Copper (Cu) centers. | Collects 4 electrons from 4 distinct Cyt c molecules and passes them simultaneously to an Oxygen ($O_2$) molecule. Oxygen is held tightly by the Copper/Iron binuclear center until it is safely reduced into 2 molecules of harmless Water ($H_2O$), preventing the formation of toxic reactive oxygen species (ROS). | 2 H⁺ |
B. Chemiosmosis & The ATP Synthase Mechanism
The continuous, relentless pumping of protons by Complexes I, III, and IV creates a massive physical and energetic discrepancy across the inner mitochondrial membrane. Because protons ($H^+$) carry a positive electrical charge and also create acidity, two separate gradients are formed simultaneously across the membrane:
- A Chemical Gradient ($\Delta$pH): The intermembrane space becomes highly acidic (low pH) compared to the relatively alkaline matrix.
- An Electrical Gradient ($\Delta \Psi$): The intermembrane space becomes highly positively charged relative to the negatively charged matrix.
Combined, these two distinct forces create the immense Proton-Motive Force (PMF). The inner membrane is strictly impermeable to ions. The only way the trapped protons can relieve this immense pressure and flow back into the matrix is through the highly specific, hollow channel provided by the enzyme ATP Synthase (Complex V).
The Binding-Change Mechanism (Rotary Catalysis)
ATP Synthase is literally a molecular nanomachine. It consists of two major structural domains:
- The $F_0$ Unit (The Rotor): Embedded entirely within the hydrophobic lipid bilayer of the inner membrane. As protons flow through its specific channel down their electrochemical gradient, they bind to specific amino acids, causing the entire $F_0$ cylinder to physically rotate within the membrane at high speeds.
- The $F_1$ Unit (The Catalytic Knob): Protrudes entirely into the mitochondrial matrix. The spinning $F_0$ rotor turns an asymmetrical central stalk (the $\gamma$ subunit) that reaches up into the center of the $F_1$ knob. As the stalk rotates, it mechanically forces the stationary catalytic subunits of the $F_1$ knob to undergo massive conformational changes, forcibly slamming ADP and $P_i$ together to forge a new ATP molecule.
Toxins, Poisons, and Uncoupling Agents
Medical entrance exams frequently test your deep understanding of respiration by asking what happens when a specific component of the machinery is maliciously blocked or chemically altered. You must firmly memorize these three distinct categories of mitochondrial interference:
-
1. Electron Transport Chain Inhibitors
Cyanide ($CN^-$) & Carbon Monoxide ($CO$): These are extremely potent, highly lethal poisons. They bind irreversibly to the Iron and Copper centers of Cytochrome c Oxidase (Complex IV) with a significantly higher binding affinity than oxygen itself.
Biochemical Consequence: The final transfer of electrons to Oxygen is completely blocked. Because the electrons have nowhere to go, the entire ETC backs up instantly (like a massive traffic jam). With electron flow stopped, no more protons can be pumped. The proton-motive force rapidly collapses to zero, and ATP synthesis halts immediately. The cell suffocates and dies of rapid energy depletion, despite the presence of abundant oxygen in the blood. -
2. ATP Synthase Inhibitors
Oligomycin: A specific macrolide antibiotic that binds directly to the $F_0$ stalk of ATP Synthase, physically blocking the proton channel like a cork jammed into a bottle.
Biochemical Consequence: Protons cannot flow back into the matrix. ATP synthesis stops immediately. However, the ETC initially continues pumping protons into the IMS until the proton gradient becomes so impossibly steep and powerful that the ETC complexes simply lack the thermodynamic energy to push any more protons against it. Eventually, the ETC seizes up and oxygen consumption halts as well. -
3. Uncoupling Agents
Mechanism: These are lipid-soluble molecules that embed themselves in the inner mitochondrial membrane and create an artificial "leaky" pore or channel. They allow protons to diffuse freely back into the matrix, completely bypassing the ATP Synthase turbine.
Biochemical Consequence: The ETC detects a catastrophic loss of the proton gradient and runs at absolute maximum speed to try and restore it. Oxygen consumption skyrockets. However, because the protons bypass the turbine, NO ATP IS PRODUCED. The massive kinetic energy of the rushing protons is instead entirely dissipated as lethal HEAT.• Synthetic Example: 2,4-Dinitrophenol (DNP). Used illicitly as an extreme, highly dangerous weight-loss drug. It burns fat stores rapidly by running metabolism at max speed, but patients literally cook to death from extreme hyperthermia from the inside out.
• Physiological Example: Thermogenin (UCP1). A natural uncoupling protein found specifically in the mitochondria of Brown Adipose Tissue (BAT). It is used safely by hibernating mammals (like bears) and human infants for "non-shivering thermogenesis" to survive extreme cold by purposefully generating heat instead of ATP.
🚀 Advanced Concept: The NADH Shuttle Systems
A critical logistical problem exists in cellular respiration: Glycolysis produces 2 molecules of NADH in the cytoplasm, but the Electron Transport Chain (which needs to oxidize that NADH to make ATP) is located inside the mitochondria. The inner mitochondrial membrane is completely and strictly impermeable to NADH. Therefore, the physical NADH molecule cannot enter. The cell solves this by using ingenious "shuttles" that extract only the high-energy electrons from the cytosolic NADH, pass them across the membrane, and hand them to a different carrier molecule inside the matrix. The type of shuttle used by a specific tissue determines the final total ATP yield of glucose.
1. Malate-Aspartate Shuttle
(High Efficiency)
- Primary Tissue Location: Heart muscle, Liver cells, Kidney cells.
- Biochemical Mechanism: Electrons from cytosolic NADH are used to reduce oxaloacetate into malate. Malate can cross the inner membrane via a specific antiport transporter. Once inside the matrix, malate is oxidized back into oxaloacetate, and the electrons are given to a resident matrix $NAD^+$ molecule, regenerating a fresh NADH inside the matrix.
- Energy Yield: Because the electrons end up on NADH (which enters the ETC at the top at Complex I), the full energy is preserved.
Yield: 2.5 ATP per cytosolic NADH.
2. Glycerol-3-Phosphate Shuttle
(High Speed)
- Primary Tissue Location: Skeletal Muscle, Brain tissue.
- Biochemical Mechanism: Electrons from cytosolic NADH reduce DHAP into Glycerol-3-Phosphate (G3P). G3P interacts with an enzyme embedded in the outer face of the inner mitochondrial membrane. This enzyme passes the electrons directly to a tightly bound FAD prosthetic group, forming FADH$_2$.
- Energy Yield: Because the electrons end up on FADH$_2$ (which bypasses Complex I and enters later at Complex II), significant energy is lost. However, this shuttle operates much faster, which is critical for muscle and brain function during sudden stress.
Yield: 1.5 ATP per cytosolic NADH.
🦠 8. Anaerobic Respiration (Fermentation)
If Oxygen ($O_2$), the essential final electron acceptor, is completely absent, a catastrophic chain reaction occurs. Complex IV cannot dispose of its electrons, so it remains stuck in a reduced state. This causes Complex III to back up, then Ubiquinone, and finally Complex I. With Complex I paralyzed, NADH cannot be oxidized back to $NAD^+$.
Why is this fatal? Step 6 of glycolysis (catalyzed by GAPDH) absolutely requires a steady, continuous supply of empty $NAD^+$ molecules to proceed. Without $NAD^+$, glycolysis halts entirely, shutting down the cell's only remaining source of ATP production, leading to rapid cell death. Fermentation serves as an emergency metabolic escape valve. Its only purpose is to provide an alternative chemical pathway to dump electrons from NADH, regenerating the $NAD^+$ required to keep glycolysis churning out a tiny, survival-level amount of 2 ATP per glucose.
A. Lactic Acid Fermentation
- Organisms & Tissues: Mammalian skeletal muscle (during intense sprinting or weightlifting when oxygen demand heavily outstrips supply), mature Red Blood Cells (which entirely lack mitochondria and thus rely 100% on fermentation forever), and Lactobacillus bacteria (used to make yogurt and cheese).
- The Reaction: A single, elegant step. Pyruvate (3C) acts as the final electron acceptor. It is directly reduced by NADH (a reaction catalyzed by the enzyme Lactate Dehydrogenase) to form Lactate (3C), also known as lactic acid. Crucially, no carbon is lost; no $CO_2$ is produced.
-
The Cori Cycle (Metabolic Burden Shifting):
Lactate accumulation in muscle tissue drastically lowers intracellular pH, interfering with contractile proteins and contributing to fatigue. The muscle cell itself cannot process the lactate further. Instead, lactate diffuses into the bloodstream and is transported to the Liver. The liver, functioning as the body's metabolic refinery, utilizes a pathway called Gluconeogenesis. It invests a massive 6 ATP to convert 2 molecules of Lactate back into 1 molecule of fresh Glucose. This newly minted glucose is then dumped back into the blood to supply the starving muscles. This cycle prevents lethal acidosis while salvaging the carbon skeleton.
B. Alcoholic Fermentation
- Organisms & Tissues: Yeast (most famously Saccharomyces cerevisiae, used in baking and brewing), numerous bacterial species, and the roots of certain plants when subjected to severe waterlogging (flooding).
- Reaction Step 1 (Decarboxylation): Pyruvate (3C) is acted upon by Pyruvate Decarboxylase. It loses a carbon atom, releasing $CO_2$ gas (this exact gas is what causes bread dough to rise and provides the carbonation in champagne and beer). The remaining 2-carbon molecule is Acetaldehyde.
- Reaction Step 2 (Reduction): Acetaldehyde acts as the final electron acceptor. It is reduced by NADH (catalyzed by Alcohol Dehydrogenase) to form Ethanol (2C), the active ingredient in alcoholic beverages.
- Crucial Distinction: Unlike lactic acid fermentation in mammals, alcoholic fermentation in yeast is strictly irreversible because the carbon atom lost as $CO_2$ gas escapes into the atmosphere and cannot be recovered to rebuild pyruvate.
🥩 9. Advanced Metabolism: Lipids & Proteins
While glucose is the standard, primary textbook model for cellular respiration, mammals rely enormously on alternative macromolecules. Fats provide the most dense, long-term energy storage, while proteins are catabolized during periods of severe starvation or extreme high-protein diets. The genius of cellular metabolism is that these diverse macromolecules do not require entirely separate pathways; instead, they are broken down and funneled directly into the existing central respiratory machinery (Glycolysis and the Krebs Cycle).
Lipid Catabolism ($\beta$-Oxidation)
- Lipolysis: Stored Triglycerides (fats) are hydrolyzed by lipases into one Glycerol backbone and three long Fatty Acid chains. The Glycerol easily enters the middle of glycolysis as DHAP.
- Activation & Transport: Fatty acids in the cytosol are chemically activated by the attachment of CoA, forming Fatty Acyl-CoA. This activation costs the equivalent of 2 ATP. Because long-chain fatty acids cannot penetrate the mitochondrial membrane, they are transported into the matrix via a highly specific transport system called the Carnitine Shuttle.
- The $\beta$-Oxidation Spiral: Once inside the mitochondrial matrix, enzymes systematically attack the fatty acid chain, cleaving off 2 carbon atoms at a time during each cycle.
Each single "cut" (cycle) yields: 1 Acetyl-CoA (which drops directly into the Krebs cycle), 1 FADH$_2$ (to the ETC), and 1 NADH (to the ETC). -
The Mathematical Power of Fat:
Palmitic Acid (16C)A 16-carbon fatty acid requires exactly 7 "cuts" to be completely degraded into 2-carbon chunks.
- Yields 8 Acetyl-CoA molecules (8 x 10 ATP via Krebs = 80 ATP).
- Yields 7 NADH (7 x 2.5 = 17.5 ATP) and 7 FADH$_2$ (7 x 1.5 = 10.5 ATP).
- Total gross ATP = 80 + 17.5 + 10.5 = 108 ATP.
- Subtract the 2 ATP used for initial activation in the cytosol.
Protein Catabolism & The Urea Cycle
Proteins are not a preferred energy source because the body does not store them for energy; every protein has a vital structural or functional role (like muscle tissue, antibodies, or enzymes). They are only catabolized when carbohydrates and fats are exhausted.
- Hydrolysis: Proteins are broken down into individual amino acids by proteases in the digestive tract or lysosomes.
- Deamination & Transamination: Before the carbon skeleton of an amino acid can be oxidized for energy, the toxic nitrogen-containing amino group ($-NH_2$) must be completely removed. This process occurs primarily in the hepatocytes (liver cells).
- The Urea Cycle (Liver): The removal of the amino group generates free ammonia ($NH_3$), which is highly toxic, especially to the brain. The liver rapidly converts this ammonia, combined with $CO_2$, into a much less toxic, highly water-soluble compound called Urea. Urea is released into the bloodstream and ultimately filtered and excreted by the kidneys in urine.
- Carbon Skeleton Entry Points: Once the nitrogen is gone, the remaining carbon structure (called an $\alpha$-keto acid) is routed into the respiratory pathway. Depending on the specific amino acid's R-group, it enters at different points:
- Glucogenic amino acids enter as Pyruvate or Krebs cycle intermediates (like $\alpha$-Ketoglutarate or Oxaloacetate) and can be used to synthesize new glucose.
- Ketogenic amino acids enter strictly as Acetyl-CoA and can only be burned for energy or converted into ketone bodies/fats.
✅ 10. Conclusion: The Final ATP Mass Balance (Per 1 Glucose)
The ultimate goal of cellular respiration is ATP synthesis. Below is the theoretical maximum yield of ATP from the complete aerobic oxidation of a single molecule of glucose. The calculations below rely on the modern, updated P/O ratios established by advanced biochemical research, which state that the energy from 1 NADH molecule pumps roughly 10 protons (yielding ~2.5 ATP), while 1 FADH$_2$ molecule pumps roughly 6 protons (yielding ~1.5 ATP).
| Metabolic Stage | Direct ATP (Substrate-Level) |
NADH Yield (x 2.5 ATP via ETC) |
FADH₂ Yield (x 1.5 ATP via ETC) |
ATP Derived from Ox-Phos | Total ATP Generated |
|---|---|---|---|---|---|
| 1. Glycolysis | 2 | 2 (Cytosolic) |
0 | 3 or 5 (Depends on Shuttle Used) |
5 or 7 |
| 2. Pyruvate Oxidation | 0 | 2 (Matrix) |
0 | 5 | 5 |
| 3. Krebs Cycle (2 Turns per Glucose) | 2 (via GTP) |
6 (Matrix) |
2 | (6 x 2.5) + (2 x 1.5) = 18 |
20 |
|
Total Theoretical Maximum Net Yield (Assuming use of the highly efficient Malate-Aspartate Shuttle in Heart/Liver cells) |
32 ATP | ||||
*Historical Note for Exams: Many older textbooks and legacy exam questions may still cite 36 or 38 ATP. This outdated number was based on earlier, rounded integer P/O estimates assuming 1 NADH yielded exactly 3 ATP, and 1 FADH₂ yielded exactly 2 ATP. Modern structural analysis of the ATP synthase turbine has definitively proven the ratios are fractional (~2.5 and ~1.5), bringing the true maximum physiological yield down to 30 or 32 ATP.
📝 Mastery Practice: IMAT Respiration Quiz
Test your comprehensive understanding of the advanced bioenergetics concepts covered in this module. These questions are meticulously styled after the rigorous, integrated logic required for the IMAT.